

51. Характеристика трёх фаз цикла Кальвина, пути превращения фосфоглицеринового альдегида.
Цикл Кальвина подразделяют на три фазы:
– карбоксилирующую РДФ + СО2 → 2ФГК;
– восстановительную ФГК → ФГА;
– регинирующую ФГА → РДФ.
Суммарный результат второй стадии: восстановление карбоксильной группы кислоты (–СООН) до альдегидной (–СНО).
Процесс превращения катализируется дегидрогеназой фосфоглициринового альдегида. Дальнейшее превращение фосфоглициринового альдегида может происходить 4 путями.
ФГА частично с помощью триозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА):
Это первый путь превращения ФГА.
Таким образом, в клетку поступают две найпростейшие формы сахаров: альдоза (ФГА) и кетоза (ФДА). Это трехуглеродные сахара (триозосахара) с присоединенной к ним фосфатной группой содержат больше химической энергии, чем ФГК. Это первые углеводы, которые образуются при фотосинтезе.
С помощью альдолазы фосфодиоксиацетон (ФДА) соединяется с другой молекулой ФГА и образуется молекула фруктозо-1,6-дифосфата (ФДФ).
Это второй путь превращения ФГА.
Фруктозо-1,6-дифосфат дефосфорилируется и превращается во фруктозо-6-фосфат (Ф-6-Ф), что сопровождается накоплением в среде неорганического фосфата. Фруктозо-6-фосфат в дальнейшем может выйти из цикла и использоваться для синтеза запасных форм углеводов: сахарозы, крахмала, других полисахаридов.
Однако ФГА (третий путь) может реагировать с эквимолярным количеством Ф-6-Ф, в результате образуются равные количества ксилулозо-5-фосфата и эритрозо-4-фосфата (транскетолаза). Затем эритрозо-4-фосфат реагирует с равным количеством ФДА и образуется седагептулозо-1,7-дифосфат (альдолаза), которая фосфорилируется до седагептулозо-7-фосфата с участием седагептулозодифосфатазы.
етвертый путь превращения ФГА связан с его реакцией с седагептулозо-7-фосфатом с образованием равных (эквимолярных) количеств рибозо-5-фосфата и ксилулозо-5-фосфата. Ксилулозо-5-фосфат эпимиризуется, а рибозо-5-фосфат изомерезуется до рибулозо-5-фосфата, последняя фосфорилируется за счет АТФ и образуется рибулозо-1,5-дифосфат – первичное соединение цикла Кальвина (акцептор СО2). В этих реакциях тратится еще три молекулы АТФ.
Из приведенных реакций цикла Кальвина видно, что фотосинтез, являясь процессом запасания энергии, тем не менее, для своего существования требует затраты энергии.
АТФ и НАДФН, которые образуются в световой стадии и используются на восстановление СО2, получили название ассимиляционной силы.
52. С2-цикл (фотодыхание), его роль в жизни растений, характеристика и локализация основных реакций.
Фотодыхание (или световое дыхание) – это индуцированное светом поглощение О2 и выделение СО2, которое наблюдается в зеленых клетках. Интенсивность фотодыхания увеличивается при повышении освещенности.
Хотя органеллой фотодыхания считается периксисома, в этом цикле непосредственное участие принимают хлоропласты и митохондрии. В клетке эти органеллы примыкают друг к другу, что также подчеркивает их функциональную зависимость. Таким образом, мы приходим к очень важному выводу: в клетке вместе с изолированностью органелл существует их интеграция.
Интенсивное фотодыхание может снизить продуктивность С3-растений на 30 %, так как при фотодыхании происходит не поглощение СО2, а его потери. Из каждых 10 молекул риболозо-1,5-дифосфата, образовавшегося в С3-цикле и включившихся в фотодыхание, одна разрушается:
53. С4-цикл (Хетча – Слэка – Карпилова), его роль в жизни растений, хар-ка и локализация основных реакций.
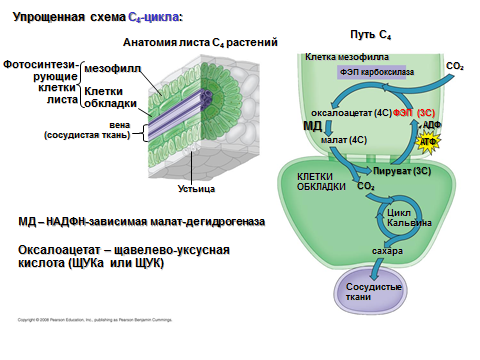
Цикл Хетча – Слэка. Цикл Кальвина – основной, но не единственный путь восстановления СО2. Так австралийские ученые М. Хетч и К. Слэк (1966) и советский ученый Ю. Карпилов (1960) выявили, что у некоторых растений, главным образом, тропических и субтропических, таких как кукуруза, сахарный тростник, сорго и другие, основная часть меченного углерода (14СО2) уже после нескольких секунд фотосинтеза обнаруживается не в фосфоглицериновой кислоте, а в щавелево-уксусной (ЩУК), яблочной (ЯК) и аспаргиновой (АК) кислотах. В этих кислотах можно обнаружить в первые секунды до 90 % поглощенного 14СО2. Через 5–10 минут метка появлялась в фосфоглицериновой кислоте, а затем в фосфоглицериновых сахарах. Так как эти органические кислоты содержат по 4 атома углерода, то такие растения начали называть С4-растениями в отличие от С3-растений, в которых радиоуглеродная метка появляется, прежде всего, в ФГК.
Это открытие положило начало серии исследований, в результате которых подробно был изучен химизм превращения углерода в фотосинтезе в С4-растениях. Акцептором углекислого газа в этих растениях является фосфоенолпировиноградная кислота (ФЕП). В настоящее время в зависимости от того, какая из органических кислот (малат или аспартат) декарбоксилируется, С4-растения делят на два типа: малатный тип (кукуруза, тростник) и аспартатный (сорго и др.) типы.
В свою очередь растения последнего типа подразделяют на две группы: растения, которые используют в реакции НАД-зависимую малатдегидрогеназу и растения, которые используют фосфоенолпируват-карбоксикиназу.
С4-растения отличаются от С3-растений и по анатомии листовой пластины. Фотосинтез идет в клетках обкладки и в клетках мезофилла. Оба типа фотосинтезирующих тканей отличаются по строению хлоропластов. Хлоропласты клеток мезофилла имеют строение, присущее большинству растений: они содержат два типа тилакоидов – тилакоиды гран и тилакоиды стромы (гранальные хлоропласты). Клетки обкладки содержат более крупные хлоропласты, часто наполненные крахмальными зернами и не имеющими гран, т. е. эти хлоропласты содержат только тилакоиды стромы (агранальные).