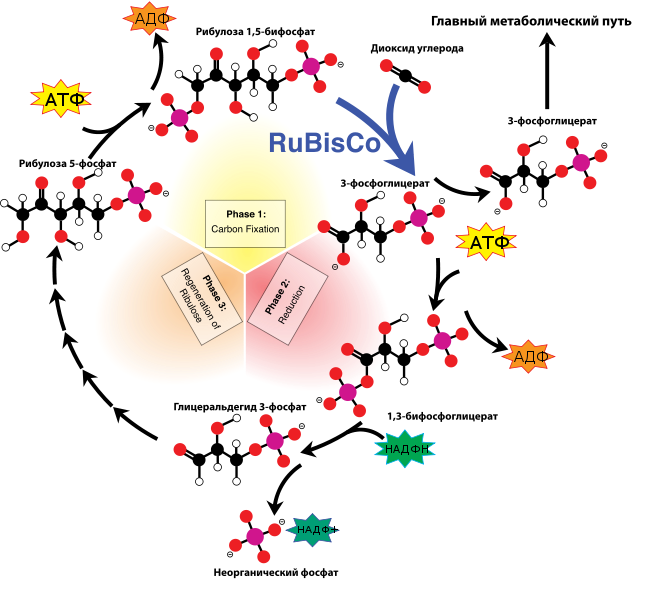
49. Роль цикла Кальвина в процессе фотосинтеза, его суммарное выражение и упрощенная схема.
Цикл Кальвина. Этот способ ассимиляции СО2 является основным и присущ всем растениям. Этот цикл начинается с присоединения СО2 к акцептору – пятиуглеродному сахару рибулозо-1,5-дифосфату (РДФ). Присоединение СО2 к тому или ионному веществу называется карбоксилированием, а фермент катализирующий такую реакцию – карбоксилазой.
В данной реакции карбоксилирование происходит с участием фермента рибулозодифосфаткарбоксилазы (РДФ-карбоксилаза). Это самый распространенный в мире фермент.
Продукт реакции, содержащий 6 атомов углерода, в присутствии воды сразу распадается на две молекулы 3-фосфоглицириновой кислоты (3-ФГК):
С данной реакции и начинается цикл Кальвина. ФГК и является, по современным взглядам, первичным продуктом ассимиляции углерода.
Для дальнейших превращений ФГК необходимы вещества световой фазы фотосинтеза: АТФ и НАДФН. Сначала 3-ФГК фосфорилируется при участии АТФ и образуется 1,3-дифосфоглицириновая кислота. Реакция катализируется ферментом фосфоглицераткиназой:
Затем происходит восстановление за счет НАДФН и образуется фосфоглицириновый альдегид ФГА:
Суммарный результат второй стадии: восстановление карбоксильной группы кислоты (–СООН) до альдегидной (–СНО).
Процесс превращения катализируется дегидрогеназой фосфоглициринового альдегида. Дальнейшее превращение фосфоглициринового альдегида может происходить 4 путями.
ФГА частично с помощью триозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА):
Таким образом, в клетку поступают две найпростейшие формы сахаров: альдоза (ФГА) и кетоза (ФДА). Это трехуглеродные сахара (триозосахара) с присоединенной к ним фосфатной группой содержат больше химической энергии, чем ФГК. Это первые углеводы, которые образуются при фотосинтезе.
Цикл Кальвина подразделяют на три фазы:
– карбоксилирующую РДФ + СО2 → 2ФГК;
– восстановительную ФГК → ФГА;
– регинирующую ФГА → РДФ.
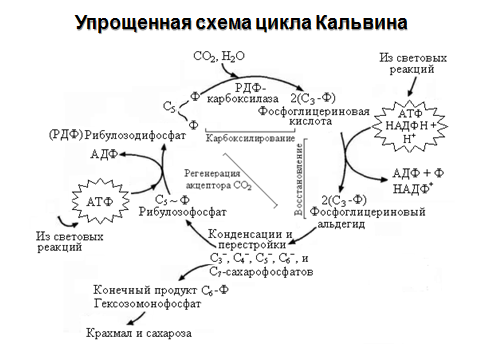
50. Химизм реакций цикла Кальвина.
цикл Кальвина — серия биохимических реакций, осуществляемая при фотосинтезе растениями (в строме хлоропластов), цианобактериями, прохлорофитами и пурпурными бактериями, а также многими бактериями-хемосинтетиками, является наиболее распространённым из механизмов автотрофной фиксации CO2.
Стадии
В цикл вовлекаются АТФ и НАДФ·Н, образованные в ЭТЦ фотосинтеза, углекислый газ и вода; основным продуктом является глицеральдегид-3-фосфат. Поскольку АТФ и НАДФ·Н могут образовываться в разных метаболических путях, цикл не следует рассматривать строго привязанным к световой фазе фотосинтеза.
Общий баланс реакций цикла можно представить уравнением:
3 CO2 + 6 НАДФ·Н + 5 H2O + 9 АТФ → C3H7O3-PO3 + 3 H+ + 6 НАДФ+ + 9 АДФ + 8 Фн + 3 H2O
Две молекулы глицеральдегид-3-фосфата используются для синтеза глюкозы.
Цикл состоит из трёх стадий: на первой под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназапроисходит присоединение CO2 к рибулозо-1,5-дифосфату и расщепление полученной гексозы на две молекулы 3-фосфоглицериновой кислоты (3-ФГК). На второй 3-ФГК восстанавливается до глицеральдегид-3-фосфата (фосфоглицеральдегида, ФГА), часть молекул которого выходит из цикла для синтеза глюкозы, а другая часть используется в третьей стадии для регенерации рибулозо-1,5-дифосфата.
Карбоксилирование
Карбоксилирование рибулозо-1,5-бисфосфата (5-углеродное соединение) осуществляется рубиско в несколько стадий. На первой кетонная группа рибулозы восстанавливается до спиртовой, между 2 и 3 атомами углерода устанавливается двойная связь. Полученное соединение нестабильно и именно оно карбоксилируется с образованием 2-карбокси-3-кето-D-арабитол-1,5-бисфосфата. Его структурный аналог 2-карбокси-D-арабитол-1,5-бисфосфат ингибирует весь процесс. Новое, уже 6-углеродное соединение, также нестабильно и распадается на две молекулы 3-фосфоглицериновой кислоты (3-фосфоглицерат, 3-ФГА).
Восстановление
Восстановление 3-фосфоглицериновой кислоты (3-ФГА) происходит в две реакции.
Сначала каждая 3-ФГА с помощью 3-фосфоглицераткиназы и с затратой одной АТФ фосфорилируется, образуя 1,3-бисфосфоглицериновая кислота (глицерат-1,3-бисфосфат).
Затем под действием глицеральдегид-1,3-фосфатдегидрогеназы бисфосфоглицериновая кислота восстанавливается НАД(Ф)·H (у растений и цианобактерий; у пурпурных и зелёных бактерий восстановителем является НАД·H) параллельно с отщеплением одного остатка фосфорной кислоты. Образуется глицеральдегид-3-фосфат (фосфоглицеральдегид, ФГА, триозофосфат). Обе реакции обратимы.
Регенерация
На последней стадии 5 молекул глицеральдегид-3-фосфатов превращаются в три молекулы рибулозо-1,5-бисфосфата.
Вначале под действием трифосфат-изомеразы глицеральдегид-3-фосфат изомеризуется в дигидроксиацетон-фосфат. Фруктозабисфосфат-альдолаза объединяет их в фруктозо-6-фосфат с отщеплением остатка фосфорной кислоты. Затем следует ряд реакций перестройки углеродных скелетов и образуется рибулозо-5-фосфат. Он фосфорилируется фосфорибулокиназой и рибулозо-1,5-бисфосфат регенерируется.
Открытие
С 1940-х гг. Мелвин Калвин работал над проблемой фотосинтеза; к 1957 с помощью CO2, меченного по углероду, выяснил химизм усвоения растениями CO2 (восстановительный карбоновый цикл Калвина) при фотосинтезе. Нобелевская премия по химии (1961).