

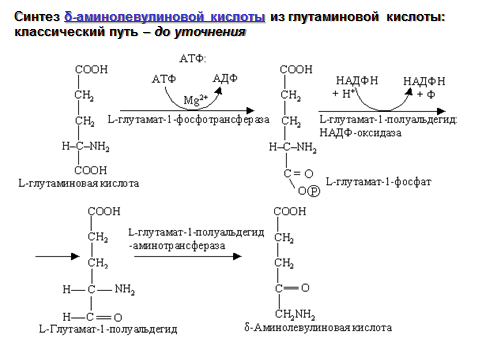
1 Стадия – синтез δ-аминолевулиновой кислоты.
субстратом служит глутаминовая кислота
δ-аминолевулиновая кислота:
δ – читается «дельта»
34. Классическая и современная схемы реакций синтеза δ-аминолевулиновой кислоты.
35. Химизм реакций синтеза хлорофилла из δ-аминолевулиновой кислоты.
1 стадия---- образуется сигма-аминолевулиновая кислота.
2 стадия – образование первичного пиррола. 2 молекулы сигма-АЛК ? порфобилиноген превращаются в присутствии фермента АЛК-дегидратазы.
3 стадия – образование циклического тетрапиррола. Четыре молекулы порфобилиногена превращаются в уропорфиноген I, а затем III.
4 стадия – происходит отщепление 4 СО2 от остатков СН2СООН -> СН3 СН3 в положениях 1, 3, 5, 8 молекулы уропорфириногена с участием фермента уропорфириногендекарбоксилазы и и образуется копропорфириноген.
5 стадия – копропорфиноген превращается в протопорфириноген 9 за счет окислительного декарбоксилирования в 2 и 4 положениях.
6 стадия –в результате реакции дегидрирования, при которой отщепляются 6Н+, из протопорфириногена 9 образуется протопорфирин.
7 стадия – происходит включение Mg (NН -> N) и образуется Mg-протопорфирин 9. Вероятно, происходит разветвление путей биосинтеза – один ведет к образованию железопорфиринов (гем), другой – к магнийпорфиринов (хлорофиллам).
8 стадия – превращение Mg-протопорфирина 9 в монометиловый эфир Mg-протопорфирина. Происходит этерификация метиловым спиртом. Далее, окисление пропионата в положении с образованием группы С=О.
9 стадия – образование протохлорофиллида. происходит в результате этирификации (этирификация – образование сложных эфиров из кислот и спиртов) карбоксильной группы метанолом, замыканием пятого (фуранового) кольца окончательным формированием боковых цепей.
10 стадия – протохлорофиллид превращается в хлорофиллид (это хлорофиллы, в молекуле которых нет терпеноидной – обычно фитольной – боковой цепи). в результате неферментативной индуцируемой светом реакции восстановления в кольце IV (появляются два атома Н)
11 стадия – образование сложного эфира хлорофиллида с фитолом, которая происходит в липидной фазе хлоропластов, поскольку фитол не растворяется в воде, с участием фермента хлорофиллаза.
36. Основные стадии и промежуточные продукты реакций синтеза каротиноидов.
Синтез каротиноидов начинается с ацетил-CоА через мевалоновую кислоту, геранилгеранилпирофосфат до ликопина. Ликопин (С40Н56) – является предшественником всех других каротиноидов. Синтез идет в темноте, но ускоряется при действии света. Ликопин имеет ациклическую структуру (отсутствие кольца с двух концов молекулы) и характерен для плодов томата. Циклизация в концах (одного или двух) молекулы ликопина приводит к образованию разных колец (β- и ε-ионовых). Так образование двух β-ионовых колец (по обоим концам молекулы) приводит к образованию β-каротина. Вообще, последовательность превращения каротиноидов следующая:
β-каротин (два β-иононовых кольца) → α-каротин (одно β-, другое ε-иононовое) → лютеин (ε-иононовые кольца и ОН-группа в кольце при С3);
β -каротин
→ зеаксантин (2 кольца β-иононовые и
группа ОН в кольцах) → виолаксантин
(два β-иононовых кольца, кроме ОН-групп
появляются эпоксигруппы в
каждом кольце).
-каротин
→ зеаксантин (2 кольца β-иононовые и
группа ОН в кольцах) → виолаксантин
(два β-иононовых кольца, кроме ОН-групп
появляются эпоксигруппы в
каждом кольце).
Фотопротекторная роль каротиноидов, т. е. защита хлорофилла от фотоокисления, объясняется их способностью взаимодействовать с возбужденными молекулами кислорода и хлорофилла. В этом случае энергия возбуждения триплетного хлорофилла и синглетного кислорода резонансным путем передается на каротиноиды, а затем рассеивается в виде тепла.
Таким образом, активный ацетил, образующийся из пировиноградной кислоты, служит исходным веществом для биосинтеза в растениях, сложных соединений к каким относятся и каротиноиды.