

7.3.3. Формирование пения птиц
Одной из моделей, успешно использованной для исследования формирования поведения и проблемы соотношения врожденного и приобретенного у разных видов птиц было становление их видоспе-цифической песни (Alcock, 1989).
Следует сразу заметить, что видоспецифическая песня развивается по-разному. Например, у зяблика (Fringilla coelebs) для развития "полной" песни необходимо, чтобы молодой самец не только прослушал ее в определенный чувствительный период, но имел бы возможность "практиковаться" в самостоятельном пении в более позднем возрасте (см. ниже). В то же время для развития песни, например, паразитических воловьих птиц (Mololrus sp.) прослушивание песни самца своего вида необязательно, молодые самцы начинают петь без соответствующего опыта в какой-либо из периодов онтогенеза.
Для выяснения вопроса о роли опыта и врожденных задатков в появлении видоспецифической песни у птиц был с успехом применен метод изолированного воспитания (депривационный эксперимент). Он показал, что не только появление видоспецифической песни птиц, но и степень сложности ее "структуры" (число колен, ритмический рисунок) в разной степени зависят от возможности слышать пение других птиц (т.е."слухового опыта"), а также от опыта общения с особями своего вида. В монографии Хайнда (1975) можно найти подробную сводку таких экспериментов, проведенных в 50– 60-е годы, т.е. в период наибольшего интереса к этой проблеме. Выяснилось, например, что птенцы серых славок (Sphenoeacus corn-munis), выращенные в изоляции от других особей своего вида в звуконепроницаемых камерах, став взрослыми, имеют в своем репертуаре все типы видоспецифических песен, причем порядок их возникновения был одинаков у всех особей.
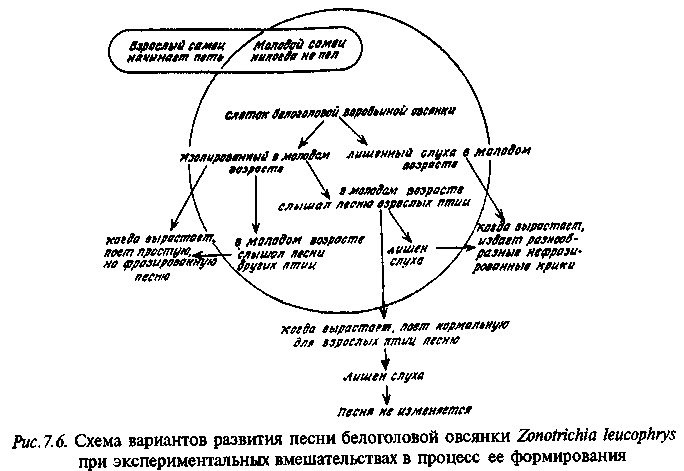
На рис. 7.6 приведена схема сложных экспериментов по влиянию раннего опыта на формирование видоспецифической песни у американской белоголовой овсянки.
7.3.4. Возможности и ограничения депривационного эксперимента
В целом данные по выращиванию животных разных видов в изоляции показали, что одни компоненты поведения развиваются в известной степени "автономно" от внешних воздействий (например, ФКД – фиксированные комплексы действий), а другие – пластичные реакции индивидуальной адаптации – могут формироваться только при наличии соответствующих раздражителей, "направляющих" развитие таких адаптации. Пример автономного развития поведения дает исследование Дж. Фентресса (Fentress, 1974) по формированию движений умывания – "груминга" – в онтогенезе мыши. Новорожденным мышатам ампутировали дистальные части передних конечностей и регистрировали на кинопленку становление реакции груминга. Несмотря на отсутствие кистей передних лап у таких животных движения предплечья и плеча в соответствующий период развития появились, однако наиболее удивительным было то, что одновременно с "умывательными" движениями конечностей появилась и реакция век в момент приближения лапы к глазу (хотя она не могла его коснуться, так как была укорочена ампутацией), он закрывался, как бы "оберегаясь" от инстинктивно ожидаемого, но несостоявшегося контакта с лапой. Этот опыт также является примером депривационного эксперимента, который доказал, что для формирования указанных движений сенсорные раздражения и опыт выполнения реакции не нужны.
Наиболее важным результатом исследований по выращиванию животных в изоляции стало понимание того, что планирование подобных опытов должно базироваться на тонком знании биологии данного вида. Планируя депривационный эксперимент, исследователь должен знать, что четкий результат он может получить только при условии выполнения ряда требований. Рассмотрим их подробно.
По своему предназначению депривационный эксперимент по зволяет лишь однозначно констатировать, что данная форма поведе ния не зависит от влияния факторов внешней среды. Если же ис следование показало, что какой-то акт поведения не проявился у животного, выращенного в изоляции, то это еще не значит, что он проявляется только в ответ на средовые воздействия и не имеет врожденной основы. Специальные исследования показали, что многие инстинктивные реакции требуют для своего "запуска" воз действия специфического для данного вида животных раздражите ля. Отсутствие такого раздражителя в условиях депривационного эксперимента может стать причиной того, что поведенческий акт не проявится, несмотря на наличие его "врожденной" основы – гото вой генетической программы развития, не требующей воздействия других факторов внешней среды (например, см.: 7.5.3).
Начиная эксперимент, исследователь должен заранее возможно более полно представлять себе весь спектр видоспецифических реак ций данного животного (этограмму). Следует обращать внимание и на патологические симптомы поведения, связанные, например, с отсутствием общения с особями своего вида. Так, в экспериментах О. Хейнрота птицы (совы, куропатки), выращенные людьми, ока зались неспособными к размножению. Это объясняется тем, что в детстве, в критический для формирования социальных привязаннос тей период у них отсутствовал опыт общения с родителями (см.: 7.1, о половом импринтинге), поэтому, став взрослыми, они не смогли опознать особей своего вида.
При проведении депривационного эксперимента необходимо концентрировать внимание на закономерностях формирования какой-то одной, заранее выбранной формы поведения, учитывая одновременно возможность разнообразных влияний на нее. Важность этого положения подтверждают результаты опытов В. Рисса (Riess, 1950). Допущенные им методические ошибки привели к неправильной трактовке данных. Рисе поставил задачу проанализировать, насколько независим от средовых воздействий комплекс движений, совершаемых крысой при постройке гнезда. Наиболее важным фактором автору представлялось лишение крыс опыта манипулирования предметами, а влияние на поведение новизны обстановки оказалось вне сферы его внимания. Рисе выращивал крыс в изоляции с 21-го дня жизни. Крысы жили в пустой проволочной клетке и питались порошкообразным кормом, так что им никогда не приходилось манипулировать предметами. Затем, когда их перевели в новую обстановку, т.е. в специальную камеру, со стенок которой свешивались бумажные полоски, ни одна из них не строила гнезда. Крысы лишь разбрасывали бумагу по клетке. Когда выращенные таким образом самки рожали детенышей, они плохо кормили их, а когда крысята расползались, плохо собирали их в гнездо (характерный элемент материнского поведения крыс), в результате чего 75% потомства погибло.
На этом основании Рисе заключил, что гнездостроительное и материнское поведение крыс было нарушено из-за отсутствия опыта манипулирования твердыми предметами.
На методическую неточность опытов Рисса указал И. Эйбл-Эйбесфельдт (см.: Lorenz, 1965), который повторил его эксперименты. В его опыте крысы были выращены так же, как и у Рисса, однако тестирование их проводили не в новой клетке, а в той, где они выросли. Все животные, у которых в клетке было постоянное место для сна (а их было около трети), и которым давали бумагу, немедленно начинали строить в этом месте гнездо (рис. 7.7).
Другие крысы пытались приступить к постройке гнезда, предварительно побродив по клетке. Автор полагает, что затруднение с выбором места для гнезда было связано с простотой и однообразием среды. Если небольшую часть клетки отделяли перегородкой, то в этом случае все воспитанные в изоляции крысы строили гнезда за перегородкой. Это позволяет заключить, что в экспериментах Рисса крысы не строили гнезд не потому, что не могли производить соответствующих действий, а потому, видимо, что боялись новой обстановки, не успевали с ней освоиться и выбрать место для сна.
Специфические фиксированные комплексы действий – гнездострои-тельные движения у крыс экспериментальных и контрольных групп были одинаковыми и, по-видимому, имели врожденный характер. Для их осуществления взрослым крысам не требуется раннего опыта манипулирования твердыми предметами, хотя исходно предполагалось, что подобный опыт необходим. Таким образом, причиной ошибочного вывода в работе Рисса был недостаточный контроль за изменением другого важного для животных внешнего фактора – степени новизны обстановки.

Депривационные эксперименты Эйбл-Эйбесфельдта показали, что в формировании некоторых других аспектов гнездостроительно-го поведения важную роль играет обучение. Если все животные охотно и сразу используют для постройки гнезда мягкую бумагу, то к использованию другого, потенциально годного для постройки гнезда материала – соломинок, – они приучаются постепенно.
Таким образом, метод изолированного воспитания, или лишение (цепривация) животного воздействий определенных внешних факторов, давший много интересной и ценной информации по развитию определенных форм поведения, необходимо использовать с учетом особенностей видоспецифического поведения животных.
7.4.
Видовая предрасположенность к некоторым видам обучения
Врожденное и приобретенное поведение с наибольшей очевидностью дифференцируются при анализе процесса обучения, особенно процесса приобретения навыков в естественных условиях жизни вида.
Этологи показали, что "приобретенное" поведение – обучение и другие виды взаимодействий со средой – имеют в ряде случаев избирательность. В настоящее время существует много данных об избирательных свойствах способности к обучению. Одним из первых на это обратил внимание У. Торп (Thorpe, 1963). В его опытах птенцы зябликов, выращенные в изоляции от сородичей, но контактировавшие с птицами других видов, из всех слышанных ими песен усваивали ту, которая более всего была похожа на песню их вида.
Интересный пример избирательной способности, "предуготовлен -ности" к обучению определенного типа, – поведение кулика-сороки. Эта птица, обитающая на побережье, по форме тела и окраске действительно напоминающая сороку, специализирована на питании моллюсками. "Инструментом" для раскалывания раковин служит характерный для птиц длинный и крепкий клюв. Уже в первый день после вылупления из яйца птенцы в более или менее точной форме выполняют соответствующие комплексы действий, характерные для этого вида при раскалывании раковины. Пока клюв птенцов не станет твердым, их кормят родители. В этот период происходит своеобразное "совершенствование" и оформление пищедобы-вательного поведения за счет следующих видов обучения: 1) образования у них условного рефлекса на моллюсков определенного вида;
тренировки способности отделять тело моллюска от раковины;
подражания родителям в способе открывания раковины – с брюшной или со спинной стороны, так как потенциально птенцы готовы к использованию обоих способов.
Используя генетические термины, можно сказать, что в генотипе этого вида зафиксирована не только "программа врожденных действий", но и "инструкция" по способам индивидуального приспособления особи. Сама возможность осуществления некоторых видов обучения, т.е. того, "что" и "как" будет выучиваться, предопределена достаточно узкой нормой реакции. Такое предопределение может быть специфично для вида, и большинство видов действительно "запрограммировано" на обучение своим собственным способом.
Был продемонстрирован также генотипический контроль общей способности к обучению. Он рассматривается в главе 8.
7.5.
Примеры развития поведения у птиц и млекопитающих