

Селективная транскрипция генов
Большинство генов не используют избирательную амплификацию в качестве механизма для обеспечения крупномасштабной экспрессии. Например, в развивающихся эритроцитах, в которых гемоглобин составляет 98% всего синтезируемого белка, не наблюдается избирательной амплификации глобиновых генов. Это значит, что определенные гены могут транскрибироваться избирательно.
Хромосомные пуфы и хромосомы типа ламповых щеток
Некоторые крупные неделящиеся клетки личинок двукрылых (таких, как Drosophila и Chironomus) содержат политенные (многонитчатые) хромосомы. В этих хромосомах репликация ДНК происходит без последующего митоза, и поэтому они содержат 512, 1024 и даже больше двойных спиральных молекул ДНК вместо одной. Клетки с политенными хромосомами никогда не делятся. Было обнаружено, что в таких хромосомах существуют «раздутые» участки. Такие участки, или пуфы, обнаруживались в различим областях хромосом из разных тканей, и их расположение менялось в ходе развития клеток. Кроме того, образование некоторых пуфов можно было индуцировать или подавить изменением физиологических условий, например при тепловой или гормональной обработке. В настоящее время известно, что эти пуфы представляют собой расплетание политенной хромосомы в менее компактную структуру, следствием этого процесса является транскрипция определенных генов.
Пуфы представляют собой расплетенную ДНК в определенных областях хромосом личинок насекомых. Так как эти хромосомы политенные, характерный вид пуфа создается тысячами нитей. Аналогичное расплетание ДНК в случае неполитенных хромосом наблюдается в ооцитах амфибий. Соответствующие структуры называют хромосомами типа ламповых щеток. На стадии диплотены мейоза компактные хромосомы амфибий образуют большие петли ДНК и втягивают их обратно после завершения этой стадии.
Транскрипция глобиновых генов
О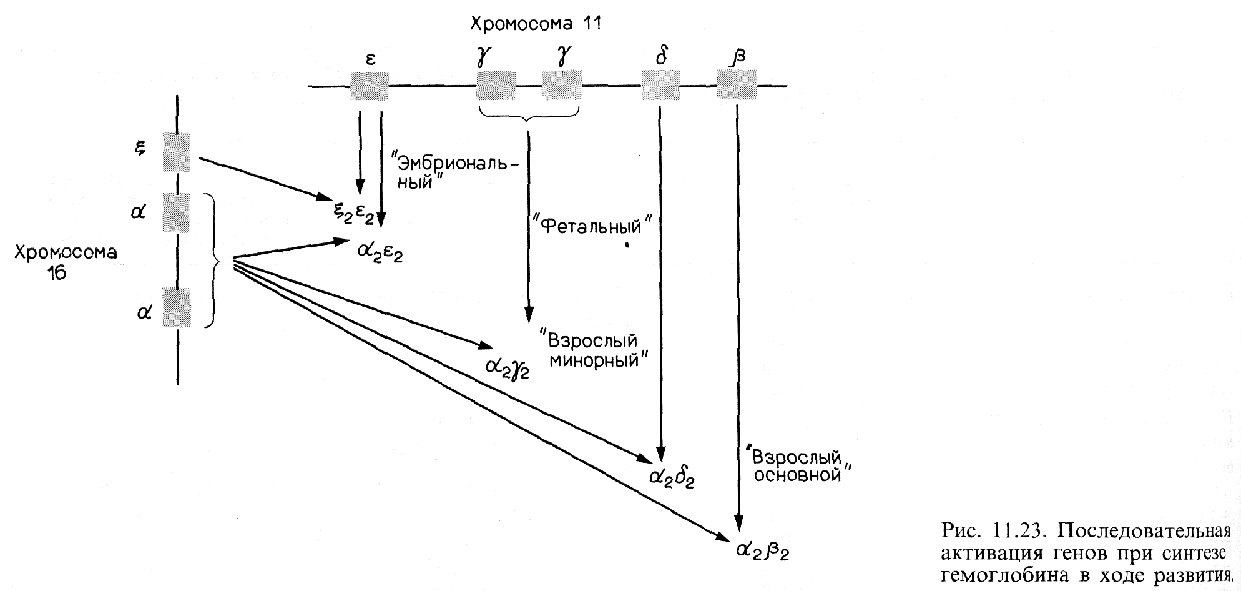 дним
из хорошо документированных случаев
регуляции дифференцировки на уровне
транскрипции является активация
глобинового гена в дифференцирующихся
эритроцитах.
У куриных зародышей и зародышей человека
в процессе развития наблюдаются изменения
в синтезе гемоглобина. На примере куриных
эритробластов можно проследить за
наиболее ранней транскрипцией гемоглобина.
В выделенной из куриного зародыша в
возрасте от 20 до 23 ч задней части темного
поля наблюдаются кровяные островки,
содержащие предшественники эритроцитов.
Зонды комплементарной ДНК к глобиновым
мРНК показывают, что эти клетки-предшественники
еще не транскрибируют глобиновые гены.
Однако после двух последующих клеточных
делений (35 ч развития) клетки (теперь
называемые эритробластами) активно
синтезируют гемоглобин. В течение этого
периода происходит включение глобиновых
генов; каким образом включаются эти
гены, остается, правда, пока неясным.
дним
из хорошо документированных случаев
регуляции дифференцировки на уровне
транскрипции является активация
глобинового гена в дифференцирующихся
эритроцитах.
У куриных зародышей и зародышей человека
в процессе развития наблюдаются изменения
в синтезе гемоглобина. На примере куриных
эритробластов можно проследить за
наиболее ранней транскрипцией гемоглобина.
В выделенной из куриного зародыша в
возрасте от 20 до 23 ч задней части темного
поля наблюдаются кровяные островки,
содержащие предшественники эритроцитов.
Зонды комплементарной ДНК к глобиновым
мРНК показывают, что эти клетки-предшественники
еще не транскрибируют глобиновые гены.
Однако после двух последующих клеточных
делений (35 ч развития) клетки (теперь
называемые эритробластами) активно
синтезируют гемоглобин. В течение этого
периода происходит включение глобиновых
генов; каким образом включаются эти
гены, остается, правда, пока неясным.
Другой тип регуляции на уровне транскрипции осуществляется на более поздних стадиях развития. У многих видов, включая курицу и человека, эмбриональный и фетальный гемоглобины отличаются от гемоглобина взрослых особей. Схематическая диаграмма типов гемоглобинов человека и генов, которые их кодируют, показана на рис. 11.23.
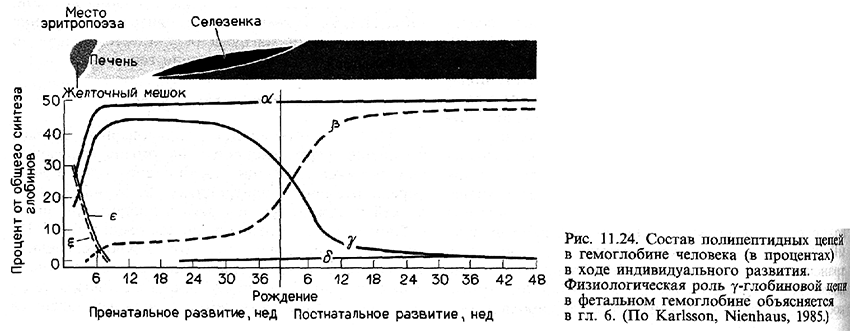
Эмбриональный гемоглобин человека состоит в оновном из двух дзета(ζ)-цепей глобина, двух эпсилон(ε)-цепей и четырех молекул гема. В течение второго месяца беременности синтез дзета- и эпсилон-цепей внезапно прекращается, тогда как синтез альфа(α) и гамма(γ)-цепей глобина усиливается (рис. 11.24). Объединение двух гамма-цепей с двумя альфа-цепями образует фетальный гемоглобин. На третьем месяце беременности активируются гены бета(β)- и дельта(δ)-цепей глобина, концентрация этих продуктов медленно возрастает, тогда как концентрация гамма-цепей постепенно снижается. Это переключение существенно ускоряется после рождения, и фетальный гемоглобин заменяется на гемоглобин взрослых, альфа2бета2. Гемоглобин взрослых людей в норме состоит на 97% из альфа2бета2, на 2-3% из альфа 2дельта2 и на 1% из альфа2-гамма2.
У человека гены дзета- и альфа-цепей глобина расположены в хромосоме 16, а гены эпсилон-, гамма-, дельта- и бета-цепей сцеплены вместе в соответствии с порядком их активации на хромосоме 11. Таким образом, представляется вероятным, что существует некий механизм, управляющий последовательным переключением генов на хромосоме 11 от генов эмбриональных глобинов к фетальным и, далее, к взрослым.
РЕЗЮМЕ Дифференциальная экспрессия генов может регулироваться на уровне транскрипции несколькими способами. Например, при гетерохроматизации значительные по размерам части хромосом могут стать генетически инертными; в случае глобиновых генов конкретный ген может быть активирован в клетке определенного типа и в определенное время. В некоторых случаях, когда требуются большие количества обычного продукта того или иного гена (как для рибосомных генов ооцитов амфибий), гены могут быть амплифицированы (для 40S-PHK) и могут синтезироваться позитивные транскрипционные факторы (TFIIIA для 5S-pPHK). Регуляция на уровне транскрипции может также определять судьбу клеток. Транскрипция таких «генов - переключателей путей дифференцировки дочерних клеток» играет существенную роль в детерминации клеток.