

Дифференциальный процессинг рнк: генерация новых белков в разных клетках в разное время
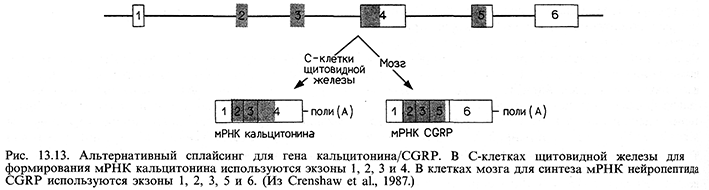
Дифференциальный процессинг РНК изучали наиболее интенсивно на примере экспрессии иммуноглобулиновых генов, однако было установлено, что он контролирует альтернативные формы экспрессии более чем 50 различных белков. В ряде случаев сплайсинг РНК может создавать в разных клетках различные белки. Процессинг определенного предшественника мРНК в некоторых клетках щитовидной железы поставляет мРНК для гормона кальцитонина. Однако в нервных клетках тот же предшественник мРНК проходит процессинг в мРНК для нейропептида CGRP. Это явление интересно само по себе, а также потому, что другой транскрипт после альтернативного сплайсинга генерирует в нейронах образование субстанции Р, а в тех клетках щитовидной железы, в которых синтезируется кальцитонин,- субстанции К. Полагают, что существуют специфичные для нейронов регулирующие сплайсинг факторы, которые функционируют на целом наборе предшественников, способных к альтернативному сплайсингу.
Полагают также, что сходным образом пять различных фибронектинов человека генерируются одним вариантом гена фибронектина. Разнообразные (и в ряде случаев органоспецифические) формы фибронектина транслируются с разных мРНК, возникающих из предшественника фибронектиновой мРНК благодаря соединению вместе различных экзонов. Альтернативный сплайсинг предшественников мРНК генерирует, очевидно, различные формы молекул адгезии нервных клеток N-MKA, а также фетальные и взрослые формы миозинов. Таким образом, альтернативный сплайсинг РНК может создавать семейство белков, кодируемых одним геном.
Регуляция экспрессии генов на уровне трансляции
После транскрипции, процессинга и выхода мРНК из ядра необходима ее трансляция, чтобы получить белок, закодированный в геноме. Регуляция на уровне трансляции представляет собой исключительно важный механизм контроля экспрессии генов. В данном случае мРНК уже образована, но она может транслироваться или не транслироваться в зависимости от определенных условий в клетке. Таким образом, контроль экспрессии генов на уровне трансляции может использоваться, когда нужен немедленный всплеск синтеза белка (как мы увидим, это происходит сразу после оплодотворения яйца) или в качестве механизма тонкой регуляции, обеспечивающей синтез строго определенного количества белка на имеющемся запасе мРНК (например, при синтезе гемоглобина). Мы узнаем также, что контроль на уровне трансляции осуществляется несколькими путями и в различных клетках используются разные пути.
Трансляция - это процесс, с помощью которого информация, содержащаяся в нуклеотидной последовательности мРНК, управляет синтезом конкретного полипептида. Этот процесс можно разделить на три этапа: инициацию, элонгацию и терминацию.
Контроль на уровне трансляции при координированном синтезе белка: продукция гемоглобина
Одной из основных проблем генетической регуляции является координированный синтез нескольких продуктов с разных участков генома. Когда развивающийся эритроцит синтезирует гемоглобин, необходимо, чтобы альфа-глобиновые цепи, бета-глобиновые цепи и молекулы гема производились соответственно в соотношении 2:2:4. Любое существенное отклонение от этого соотношения приводит к тяжелым заболеваниям.
Результаты недавних исследований показали, что пропорциональный синтез компонентов гемоглобина регулирует молекула гема. Достигается это двояким образом. Во-первых, избыток гема будет выключать свой собственный синтез. Осуществляется выключение синтеза посредством инактивации 8-аминолеву-линатсинтазы (DALA-синтазы), первого фермента на пути продукции гема. Таким образом, когда количество гема превосходит количество молекул, способных присоединить его, дальнейшая продукция гема прекращается. Во-вторых, избыток гема стимулирует синтез глобинов. Трансляционный контроль синтеза глобинов не ограничивается этим. В диплоидной клетке имеются четыре активных альфа-глобиновых гена и лишь два активных бета-глобиновых гена. Если бы каждый ген транскрибировался и транслировался с одинаковой скоростью, то следовало ожидать, что количество альфа-глобиновых молекул вдвое превысит количество бета-глобиновых молекул. Этого, естественно, не происходит. Обнаруживается отношение 1,4:1 для а-мРНК:в-мРНК, но 1:1 для белков. Таким образом, уравнивание количеств белков обусловлено регуляцией на уровне трансляции. Таким образом, правильное соотношение альфа-глобина, бета-глобина и гема устанавливается на стадии инициации трансляции. Итак, синтез гемоглобина подвержен регуляции на уровнях транскрипции и процессинга РНК, однако итоговая молекула формируется при тонкой координации на уровне трансляции.