

Коллоквиум 6.
1. Гормоны - гликопротеины: ЛГ, ФСГ, ТТГ. Место образования – гипофиз. Действуют на 7-TMS рецепторы G-белок ФЛ С расщепляет фосфатидилинозитол на ИФ3 или ДАГ или 7-TMS-рецепторы G-белок активация аденилатциклазы цАМФ Протеинкиназа А фосфорилирование белков.
2. Гормоны – простые белки:
а) пептиды (< 20 АК): окситоцин, вазопрессин, тиреотропин- и гонадотропинрилизинг гормоны, соматостатин – гипоталамус
б) белки (> 20 АК): кортикотропин- и соматотропинрилизинг гормоны – гипоталамус, пролактин, СТГ, АКТГ – передняя доля гипофиза, инсулин, глюкагон – островки поджелудочной железы, холецистокинин, панкреозимин – энтероэндокринная система, паратирин – паращитовидная железа, кальцитонин – С-кл щитовидной железы.
Пример действия простого белка: вазопрессин + 7-TMS рецептор Gq-белок ФЛ С ДАГ + ИФ3 Ca2+ 1) вазоконстрикция 2) усиливает реабсорбцию воды в нефроне.
3. Гормоны – производные АК: мелатонин – эпифиз, катехоламины (А, НА, дофамин) – нейроны и мозговое вещество надпочечников, Т3 и Т4 (тироксин) – щитовидная железа.
Т3 + ядерный рецептор (есть цинковые пальцы, но нет HSP) димеризация рецептора Т3 с рецептором ретиноевой кислоты активный гетеродимер связь с промотором ДНК активация транскрипции.
4. Особенности биологического действия гормонов:
1) находятся в очень низкой концентрации в крови (10-6-10-12 М)
2) обязательна связь с рецептором, включающая каскадный механизм и усиление гормонального сигнала (на поверхности клетки, в цитоплазме или ядре)
3) изменение скорости синтеза ферментов и их активности
4) регуляция секреции по принципу прямой или обратной связи
5. Классификация рецепторов:
1. Мембранные рецепторы
а) 7-TMS рецепторы, взаимодействующие с гетеротримерными G-белками
б) 1-TMS рецепторы со свойствами гуанилатциклазы, тирозинкиназы, протеинфосфатаз, сер/тре протеинкиназ, взаимодействующие с тирозинкиназой.
в) ионные каналы (лиганд-зависимые, потенциал-зависимые, щелевые контакты)
2. Ядерные (со свойствами регуляторов транскрипции) и цитозольные рецепторы.
Функциональное назначение G-белка: 1) Gsa – активация АЦ, 2) Gia – ингибируют АЦ, 3) Gqa – реагируют с ФЛ С и активируют ее 4) Goa – открывают Ca2+-каналы.
6. Вторичные посредники в проведении гормонального сигнала в клетку: а) циклические мононуклеотиды (цАМФ, цГМФ) б) ИФ3 в) ДАГ г) другие липиды д) цитозольный Ca2+ е) оксид азота.
7. Механизмы образования и роль вторичных посредников в механизме действия гормонов:
1) Г + 1-TMS рецептор гуанилатциклазу цГМФ активация цГМФ-зависимой протеинкиназы G фосфорилирование остатков серина и треонина в белках-мишенях.
2) Г + 7-TMS рецептор Gs- или Gi-белок АЦ или АЦ цАМФ или цАМФ. цАМФ активация протеинкиназы А фосфорилирование белков-мишеней.
3) Г + 7-ТМS рецептор Gq-белок активация ФЛ С расщепление фосфатидилинозитола на ДАГ и ИФ3. ДАГ активация протеинкиназы С фосфорилирование белков-мишеней. ИФ3 открытие Ca2+-каналов Ca2+ непосредственные эффекты (участие в сокращении и т.п.) или в комплексе к белками (кальмодулином)
4) Г + 7-TMS рецептор Goa открытие Ca2+-каналов Ca2+.
8. Механизм передачи гормонального сигнала к фосфолипазе С:
Г + 7-TMS рецептор Gq-белок активация ФЛ С расщепление фосфатидилинозитола на ДАГ и ИФ3. ДАГ активация протеинкиназы С фосфорилирование белков-мишеней. ИФ3 открытие Ca2+-каналов Ca2+ непосредственные эффекты (участие в сокращении и т.п.) или в комплексе с белками (кальмодулином).
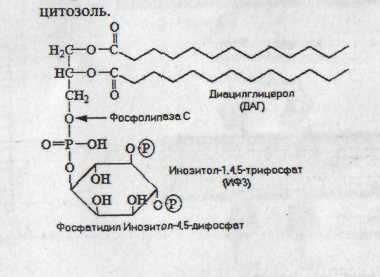
9. Роль ионов Ca2+: 1) являются вторичными посредниками, участвуя в амплификации первичного сигнала (гормона). 2) могут оказывать прямое активирующее действие (активация протеинкиназы С) или опосредованное через специальные Ca2+-связывающие белки (кальмодулин). Комплекс Ca2+-кальмодулин активирует ряд ферментов (аденилатциклазу).
10. Активация мембранносвязанной гуанилатциклазы: атриопептид, пептиды, секретируемые яичником, эндотоксин бактерий + 1-TMS рецептор изменение конформации передается на подмембранный домен (мембранносвязанную гуанилатциклазу) измененине ее конформации активация гуанилатциклазы из ГТФ образуется цГМФ активация фосфодиэстеразы из цАМФ образуется АМФ.
Активация растворимой гуанилатциклазы: NO(образуется из аргинина с помощью оксигеназных реакций) + растворимая гуанилатциклаза активация гуанилатциклазы (за счет изменения структуры гема, входящего в состав гуанилатциклазы) образование цГМФ из ГТФ активация протеинкиназы G а) дефосфорилирование белков легких цепей миозина расслабление ГМК сосудов вазодилятация или б) фосфорилирование СЕР или ТРЕ.
11. Механизм усиления – амплификация – активация одним ферментом других ферментов и увеличение числа вовлекаемых молекул в геометрической прогрессии в ферментных каскадах.
12. Общие принцип синтеза гормонов пептидной природы: синтез препрогормона на грЭПС по механизму матричного синтеза удаление сигнального пептида и превращение препрогормона в прогормон транспорт прогормона в КГ удаление дополнительных пептидов прогормона и образование гормона упаковка в секреторные гранулы для последующей секреции.
13. Гормоны гипоталамуса белково-пептидной природы. В зависимости от того, вызывают освобождение гормонов гипофиза или угнетают их выделение называются либеринами или статинами. Семь либеринов: кортиколиберин, тиролиберин, люлиберин, фоллиберин, соматолиберин, пролактолиберин, меланолиберин и три статина: пролактостатин, меланостатин, соматостатин. Регуляция по принципу обратной связи (в основном) или прямой (взаимовлияние ЛГ и эстрогена).
14. См. в. 13. Механизм действия: Г + 7- TMS рецептор Gq-белок ФЛ С ИФ3 Ca2+ высвобождение гипофизарного гормона-тропина.
15. Место образования вазопрессина и окситоцина: нейроны гипоталамуса, затем транспортируются в заднюю долю гипофиза в комплексе с белками-нейрофизинами I, II и III. По химической природе пептиды с дисульфидной связью. Механизм действия: окситоцин + 7-TMS рецептор Gq-белок ФЛ С ДАГ + ИФ3 Ca2+ 1) выделения молока 2) сокращения матки 3) перистальтики семявыносящих канальцев.
Вазопрессин: вазопрессин + 7-TMS рецептор Gq-белок ФЛ С ДАГ + ИФ3 Ca2+ 1) вазоконстрикция 2) усиливает реабсорбцию воды в нефроне.
16. Путь передачи гормонального сигнала от рецептора гормона роста: СТГ + 1-TMS рецептор димеризация р-ра гомодимер + JAK-белки (обладают тирозинкиназной активностью):
а) JAK-2-киназа STAT-белки (переносчики сигнала и активаторы транскрипции) связь с промотором активация транскрипции
б) активация IGF (инсулиноподобный фактор роста-I).
Основные эффекты СТГ: рост костей, хрящей, мышц, синтез белков, НК, липолиз, глюконеогенез, лактогенная активность, уровень гл в плазме.
17. Гормоны щитовидной железы:
1) производные АК а) T3 (3,5,3’-трийод-L-тиронин) б) Т4 (3,5,3’,5’-тетрайод-L-тиронин или тироксин)
2) пептид - кальцитонин
Т3 + ядерный рецептор (есть цинковые пальцы, но нет HSP) димеризация рецептора Т3 с рецептором ретиноевой кислоты активный гетеродимер связь с промотором ДНК активация транскрипции.
Основные эффекты Т3 и Т4:
1) влияет на нормальное развитие плода 2) синтез белков (в первую очередь энергетического обмена) 3) потребление О2 и теплопродукцию за счет увеличения Na+/К+-АТФазы во всех тканях, кроме мозга, селезенки, яичек 4) синтез эритропоэтина и содержание 2,3-ДФГ в эритроцитах 5) усиливает основной обмен 6) активность супероксиддисмуттаза 7) глюконеогенез, усвоение гл, гликогенез, липолиз, гликогенолиз 8) число рецепторов к ЛПНП 9) сердце: транскрипцию легкой цепи миозина, Ca2+-АТФазу, число β-Ар 10) стимулирует дыхательный центр 11) повышение моторики ЖКТ 12) повышает резорбцию костей.
Основные эффекты кальцитонина: резорбции Ca2+ и РО43- из костей, реабсорбцию Ca2+ и РО43- в почках.
18. Гормон паращитовидной железы – паратирин – пептид. Механизм действия: 7-TMS р-р Gs-белок АЦ цАМФ ПК А фосфорилирование белков-эффекторов. Основные эффекты: уровень Ca2+ и РО43-, выделение Ca2+ и (РО43-) из костей, остеолиз остеокластами, реабсорбцию Ca2+ почками и секрецию РО43-, образование витамина D, абсорбцию Ca2+ из ЖКТ.
19. Основные регуляторы обмена кальция в организме: паратирин (см. в. 18), кальцитонин (см. в. 17), витамин D3 (1,25 – (ОН)2- дигидроксихолекальциферол) – стероид, вырабатывается в почках. Действие витамина D3: 1) уровня Ca2+ и РО43- 2) абсорбции Ca2+ и РО43- из ЖКТ 3) реабсорбцию кальция из костей и поддерживает функционирование систем переноса Ca2+.
20. Глюкагон – пептид, образуется α-клетками островков Лангерганса поджелудочной железы. Механизм действия (7-TMS р-ры): гликогенолиз, глюконеогенез, образование кетоновых тел и липолиз.
21. См. в. 3 22. См. в. 17. Этапы синтеза тиреоидных гормонов:
1) захват иодида I- из плазмы в тироциты с помощью насосов базальной мембраны Na+/I- и Na+/K+-АТФазы
2) окисление и органификация иодида в тироцитах с помощью тиреопероксидазы: I-I. и включение его в состав тиреоглобулина
3) экзоцитоз тиреоглобулина в просвет фолликула
4) иодирование тирозиновых остатков тиреоглобулина в полости фолликула: 3-монойодтиронин (MIT) + MIT DIT (3,5-дийодтиронин), DIT + DIT T4, MID + DIT T3 (очень редко, обычно Т3 образуется из Т4 с помощью дейодазы)
5) апикальная мембрана (белок пендрин) в виде лепестков розы охватывает капли коллоида и поглощает их с образованием лизосом, затем Т3 и Т4 попадают в кровь, где переносятся белками к мишеням.
23. Гормоны мозгового слоя надпочечников: катехоламины (адреналин, норадреналин). Химическая природа: производные АК. Механизм действия (цАМФ):1) секрецию глюкагона и секрецию инсулина 2) секрецию паратирина 3) гликогенолиз 4) глюконеогенез в печени 5) липолиз.
24. Принципы синтеза стероидных гормонов (половых, глюко- и минералкортикоидов):
1) предшественником синтеза стероидов является холестерол
2) в генотипе нет информации для синтеза непосредственно стероидных гормонов, а содержится только информация о белках-ферментах, участвующих в их синтезе
3) стероидные гормоны не запасаются
4) поступают в кровоток по мере синтеза
25. Схема синтеза стероидных гормонов:
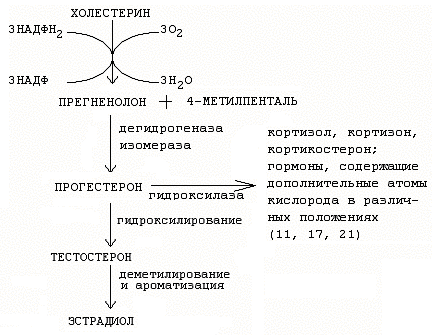
26. Гормоны половых желез: женские (эстроген, прогестерон, активин, ингибин, фолистатин, релаксин – синтезируются в яичниках), мужские (андрогены – тестостерон и дигидрокситестостерон, ингибин, активин – синтезируются в семенниках). Гормоны стероидной природы: эстроген, прогестерон, тестостерон, белковой природы: активин, фолистатин, ингибин, релаксин.
Механизм действия стероидных половых гормонов, кроме эстрогенов: связываются с рецептором в цитоплазме, затем комплекс гормон-рецептор попадает в ядро. Непосредственно в ядре располагается рецептор эстрогенов.
Эффект эстрогенов: 1) пролиферация эндотелия 2) возбудимость миометрия 3) рост протоков молочной железы 4) секреция пролактина (но не действие) 5) транспортные белки печени 6) анаболики 7) приводят к гипотермии 8) задерживают воду.
Функции прогестерона: 1) развитие желез матки, индуцирует превращение клеток стромы в децидуальные клетки 2) возбудимость миометрия 3) рост альвеол грудной железы 4) катаболик 5) температуру тела 6) натрийуретик
Эффект андрогенов: 1) сперматогенез 2) вторичные половые признаки 3) анаболик
27. Глюкокортикоиды – гормоны стероидной природы: кортизол, кортикостерон. Рецепторы расположены в цитоплазме, состоят из трех частей:
1) домен АF1 – активатор транскрипции 2) ДНК-связывающий домен (имеет цинковые пальцы, закрытые белком – ингибитором hsp) 3) гидрофобный карман.
Гормон + гидрофобный карман сброс белка теплового шока (hsp) освобождение цинковых пальцев димеризация рецепторов (гомодимер) связь с ДНК запуск транскрипции.
Действие кортизола: 1) глюконеогенез 2) липолиз и протеолиз 3) усвоение глюкозы 4) иммунный ответ 5) рост костей 6) сурфактант 7) ферменты ЖКТ 8) модулирует ЦНС.
28. Минералкортикоиды: альдостерон – стероидной природы. Действие альдостерона: 1) реабсорбция натрия почками 2) секреции К+ почками. Рецепторы располагаются в цитоплазме.
29. Рецептор инсулина: гликопротеин, 1-TMS-рецептор на поверхности клеток-мишеней. Гетеродимер, состоит из двух субъединиц – α и β. Инсулин связывается с α-субъединицей, β-субъединица преобразует полученный сигнал (обладает тирозинкиназной активностью и содержит участок аутофосфорилирования).
Г + рецептор изменение конформации α-субъединиц активация тирозинкиназной активности β-субъединицы активация инсулинорецепторного субстрата (IRS) запуск:
а) фосфатидилинозитолкиназный путь ДАГ + ИФ3 активация 50 ферметов обмена углеводов, липидов, белков, активация движения GLUT4.
б) МАП-киназный путь (митогенактивируемая протеинкиназа) факторов транскрипции синтез белков, стимуляция деления кл.
в) активация RAS-белка, связанного с рецептором фактора роста.
30, 31. Влияние инсулина на метаболизм углеводов:
1) на транспорт глюкозы через мембрану: облегчает диффузию гл в клетки, кроме печеночных за счет увеличения числа переносчиков GLUT4. В печеночных кл: индукция глюкокиназы гл в гл-6-ф концентрация свободной гл поддерживается на низком уровне гл проникает в кл путем простой диффузии по градиенту концентрации.
2) на утилизацию гл: усиливает гликогенез (например, повышает активность гликогенсинтазы)
3) на образование гл: ингибирует глюконеогенез (ингибирует синтез ФЕПКК).
4) на метаболизм гл: содержание гл в крови.
Расстройство метаболизма углеводов при диабете: 1) гипергликемия из-за а) пониженного проникновения гл в клетки б) утилизации гл различными тканями в) глюконеогенеза в печени 2) снижение гликогенеза.
32. Влияние инсулина на метаболизм липидов:
1) стимулирует липогенез в жировой ткани (за счет притока ацетил~КоА и НАДФН для синтеза ВЖК, поддержания нормального уровня ацетил~КоА-карбоксилазы, притока глицерола) 2) мощный ингибитор липолиза в печени и жировой ткани (ингибирование гормон-чувствительной липазы)
Расстройство метаболизма углеводов при диабете: повышенный липолиз содержание в плазме СЖК кетогенез, кетонурия, кетонемия .
33. Влияние инсулина на метаболизм белков: усвоение белка, усиливая утилизацию АК, стимулирует синтез, уменьшает распад (на уровне трансляции мРНК).
Нарушение при сахарном диабете: повышенный катаболизм белков повышенное содержание АК в плазме, потеря азота с мочой.
34. Восстановительный путь обмена глюкозы при сахарном диабете – образование сорбитола: глюкоза проникает в клетку сорбитол (с помощью альдозоредуктазы и НАДФН+Н+). Это наиболее интенсивно протекает в артериальных стенках, кл Шванна и т.д. Сорбитол вызывает осмотическое набухание клеток и нарушение их функций.
Основной механизм повреждения тканей при сахарном диабете - гликозилирование белков - не ферментативная реакция глюкозы со свободными аминогруппами белковой молекулы (лиз, арг). При этом нарушаются функции белков в результате изменения заряда белковой молекулы, ее конформации или блокирования активного центра, что приводит со временем к ангиопатиям (повреждениям базальных мембран сосудов).
Недостаток инсулина мобилизация жиров заполнение кровеносного русла СЖК и ТАГ ингибирование синтеза ВЖК катаболические пути не справляются с избытком образующегося ацетил-КoA в печени из ацетил-КoA синтезируются кетоновые тела накопление в крови (кетоз), т.к. скорость их образования превышает возможности клеток по их использованию метаболический ацидоз.
35. Метаболизм глюкозы в инсулиннезависимых тканях:
1) глюкоза проникает в клетку сорбитол (с помощью альдозоредуктазы и НАДФН+Н+)фруктоза (под действием сорбитолДГ).
В норме этот путь происходит в клетках артериальных стенок, клетках Шванна, в эритроцитах, в хрусталике и сетчатке глаза, в семенниках
2) глюкоза связывается с аминогруппами белков (гликозилирование), что изменяет строение белков и нарушает их функции. Может быть гликозилирование гемоглобина, белков базальных мембран изменение капилляров, утолщение базальных мембран.
36. Биохимическая диагностика сахарного диабета:
1) качественное и количественное двукратное определение гл в крови по глюкозооксидазному методу.
2) тест на толерантность к глюкозой (пробу с нагрузкой глюкозой).
3) метод Хагедорна-Йенсена - количественный метод определения уровня сахара в крови, основанный на способности сахаров восстанавливать в щелочной среде красную кровяную соль в желтую; о содержании сахара судят по избытку красной кровяной соли, определяемому титрометрией.
4) качественное и количественное определение сахара в мочи (качественные реакции: Тромера, Фелинга, количественные реакции: глюкозооксидазный метод).
37. Клинико-диагностические значения оценки гликемических кривых:
1) у больных с разными формами диабета: нарастание гликемической кривой происходит медленнее, в большинстве случаев отмечается глюкозурия. Чем тяжелее заболевание, тем позже достигается максимум гликемии и тем он выше. Понижение кривой происходит очень медленно, чаще оно растягивается на 3-4 ч.
2) гиперфункция щитовидной железы: гликемические кривые с более быстрым, чем в норме, подъемом, что вызвано более интенсивным обменом веществ и возбуждением симпатического отдела ВНС
3) аденома островков Лангерганса, гипотиреоз (микседема), болезнь Аддисона: низкий исходный уровень кривой, низкая ее вершина и высокий постгликемический коэффициент.
38. Функции печени:
1) гомеостатическая 2) метаболическая (в обмене белков, липидов, углеводов) 3) экскреторная 4) антитоксическая 5) обмен желчных пигментов.
39. Роль печени в углеводном обмене – поддержание постоянства глюкозы в крови:
а) набором ферментов синтеза и распада гликогена (гексокиназа, глюкокиназа, гликогенсинтаза, гл-6-фосфатаза)
б) набором ферментов глюконеогенеза
в) набором ферментов, превращающих другие гексозы (галактозу, фруктозу) в глюкозу.
40. Функциональные пробы, характеризующие роль печени в углеводном обмене:
а) нагрузочная проба галактозой – в норме выведение галактозы в моче не превышает 8% через 12 ч.
б) нагрузка глюкозой.
41. Роль печени в обмене липидов (их депонировании):
а) синтез, распад, удлинение, укорочение ЖК
б) распад, синтез и модификация СЖК
в) синтез большинства ЛПНП и 90% ХС в организме
г) из ХС синтезируются желчные кислоты, входящие в состав желчи.
42. Роль печени в обмене белков: АК используются для
а) синтеза белков плазмы крови (альбуминов, глобулинов, факторов свертывания – протромбин, фибриноген, факторы V, VII, IX, X, XI, XII, ферментов (липопротеинлипаза, холинэстераза)
б) образования α-кетокислот
в) глюконеогенеза из глюкогенных АК
г) кетогенеза из кетогенных АК
д) получения энергии (в ЦТК)
е) синтез креатина, холина, мочевой кислоты.
43. Роль печени в азотистом обмене: см. в. 42
1) Аммиак, образующийся в реакциях метаболизма в печени и возникающий в процессе гниения белков в толстом кишечнике, превращается в гепатоцитах в мочевину и обезвреживается.
2) В печени синтезируется креатин, затем он поставляется в кровь для использования в дальнейшем скелетной и сердечной мышцами.
Синтез креатина: 1) глицин + аргинин орнитин + гуанидиноацетат (под действием аргинилглицинтрансаминазы) 2) гуанидиноацетат + S-аденозилметионин аденозилгомоцистеин + креатин (под действием гуанидиноацетаттрансметилазы). Креатин + АТФ креатинфосфат (под действием креатинфосфокиназы) – депо энергии в мышцах.
Снижение уровня мочевины в крови наблюдается при заболеваниях: 1) патологическом изменении значительной части паренхимы печени 2) недостаточности белка в питании 3) продолжительном голодании 4) врожденном нарушении нормального протекания цикла мочевины (у детей).
44. Функциональные пробы, характеризующие белоксинтезирующую функцию печени:
Т.к. в печени синтезируются основные белки крови – альбумины и глобулины, на основании определения соотношения этих белков в плазме крови можно судить о белок-синтезирующей функции печени.
Альбумин-глобулиновый коэффициент А/Г в норме 1,5-1,7. При острых поражениях печеночной паренхимы A/Г за счет уменьшения уровня альбуминов падает осмотическое давление крови развитие периферических отеков. Т.к. синтез -глобулинов при этом не нарушается, повышается их концентрация относительно альбуминов и наблюдается дислипопротеинемия.
45,46. Печень играет исключительную роль в обезвреживании как поступающих в организм ксенобиотиков, так и образующихся в организме токсичных и непригодных для дальнейших превращений продуктов метаболизма.
Антитоксическая функция печени – обезвреживание ксенобиотиков, продуктов распада БАВ, сильнодействующих физиологических веществ, нормальных метаболитов и лекарств следующими способами:
а) метилирование (вит PP N-метилникотинамид выводится из организма)
б) ацетилирование (сульфаниламидные препараты)
в) окисление (алкоголь, ароматические углеводороды, катехоламины, биогенные амины)
г) восстановление (нитробензол)
д) синтез мочевины (аммиак)
е) коньюгация с сильнополярными отрицательно заряженными молекулами (активными формами глюкуроновой или серной кислот – УДФ-глюкуроновой кислотой и ФАФС) (стероидные гормоны, билирубин, продукты гниения белков, фенолы, желчные кислоты)
ж) окисление микросомными комплексами ферментов – превращают гидрофобные соединения в гидрофильные. С помощью монооксигеназ (НАДФ-зависимые с цитохромом P450) окисляются лекарства и яды, путем восстановления с участием НАДН-зависимых реакций могут быть инактивированы дисульфидные и нитрогруппы кетонов, альдегидов.
з) окисление пероксисомными ферментами (оксидазой мочевой кислоты, лактооксидазы, каталазы)
и) с помощью цистеин-богатого металлотиопротеина (связывание и обезвреживание тяжелых металлов Cd2+, Cu2+, Hg2+, Zn2+).