

6.2.3. Третинна структура
Під третинною структурою білків і нуклеїнових кислот звичайно розуміють спосіб просторової укладки поліпептидних і полінуклеотидних ланцюгів в 3-вимірному об’ємі. Відповідь на питання, як організована третинна структура біополімерів, вимагає застосування прецизійних експериментальних методів дослідження (зокрема, методів рентгеноструктурного аналізу, електронної мікроскопії тощо) та сучасних комп’ютерних технологій обробки цих експериментальних даних.
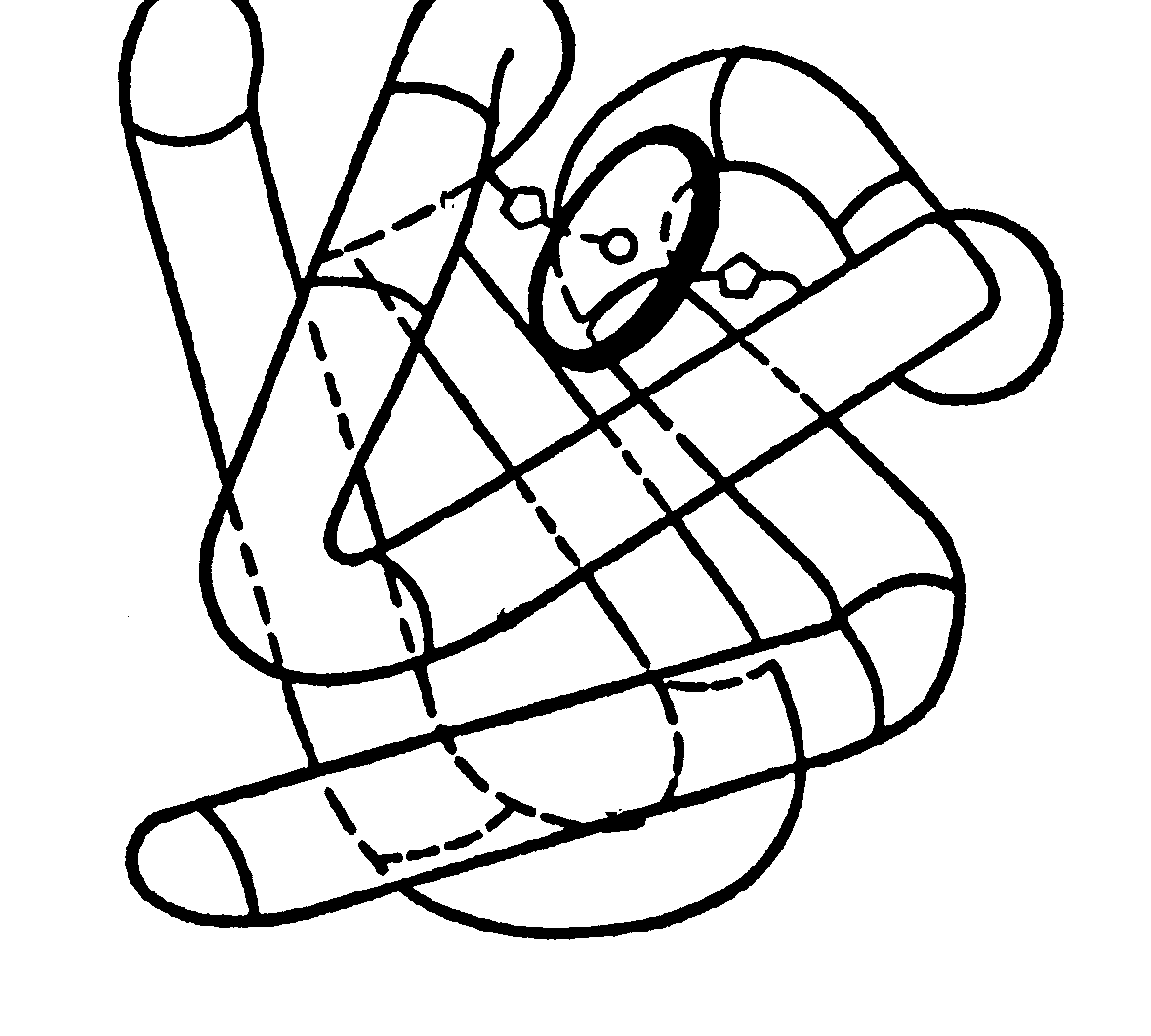
Вперше третинна структура білкової молекули була визначена Дж. Кендрю для міоглобіна кита-кашалота. Цей білок, що містить в собі 153 амінокислотних залишків, відповідає за перенос кисню в м’язах. Виявилося, що третинна структура поліпептидного ланцюга міоглобіну має вигляд скрученої трубки, що досить щільно укладена навколо гема (мал. 6.10).
|
|
Мал. 6.10. Третинна структура білкової молекули міоглобіну. |
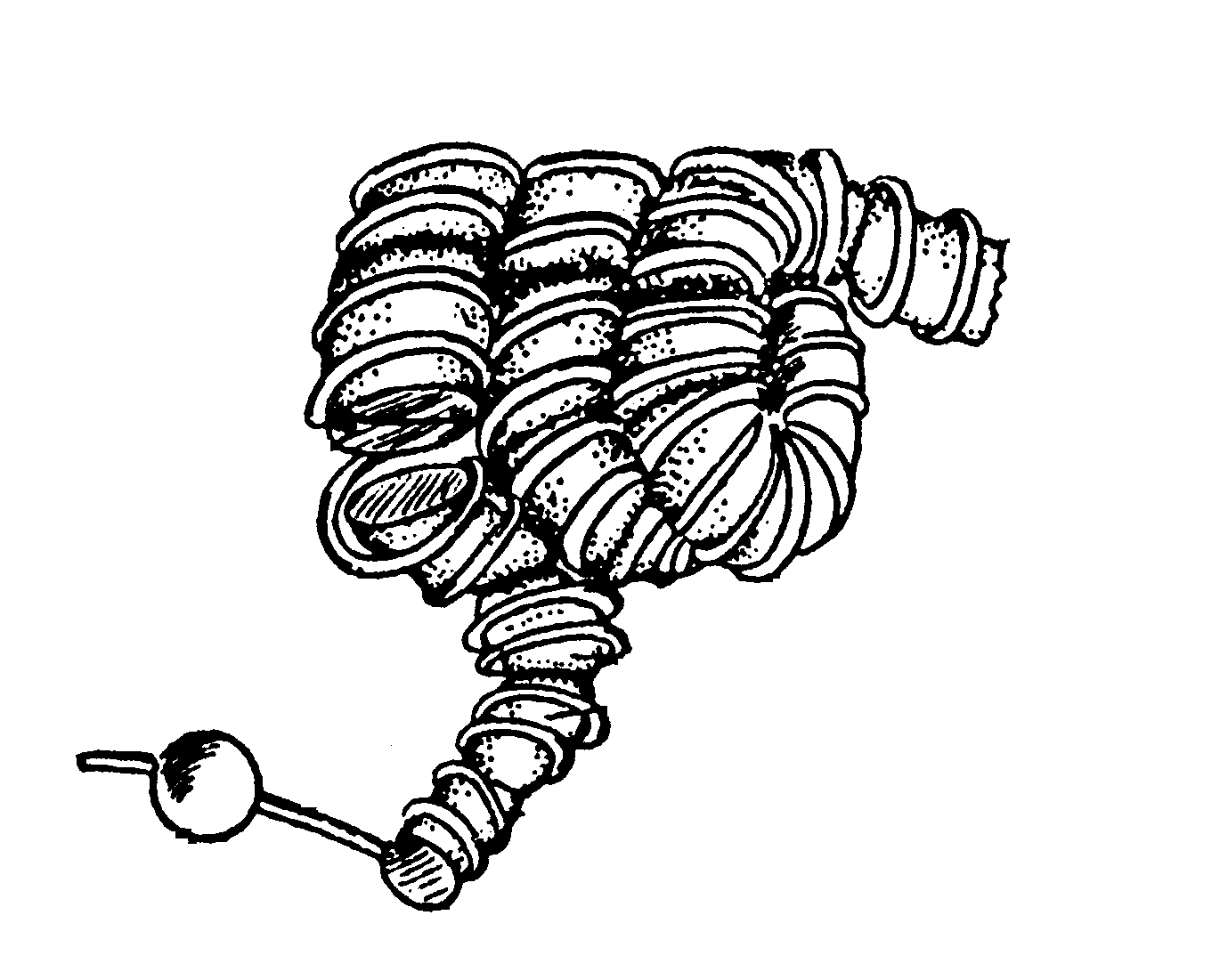
Мал. 6.11. Третинна структура ДНК в еукаріотичних клітинах. |
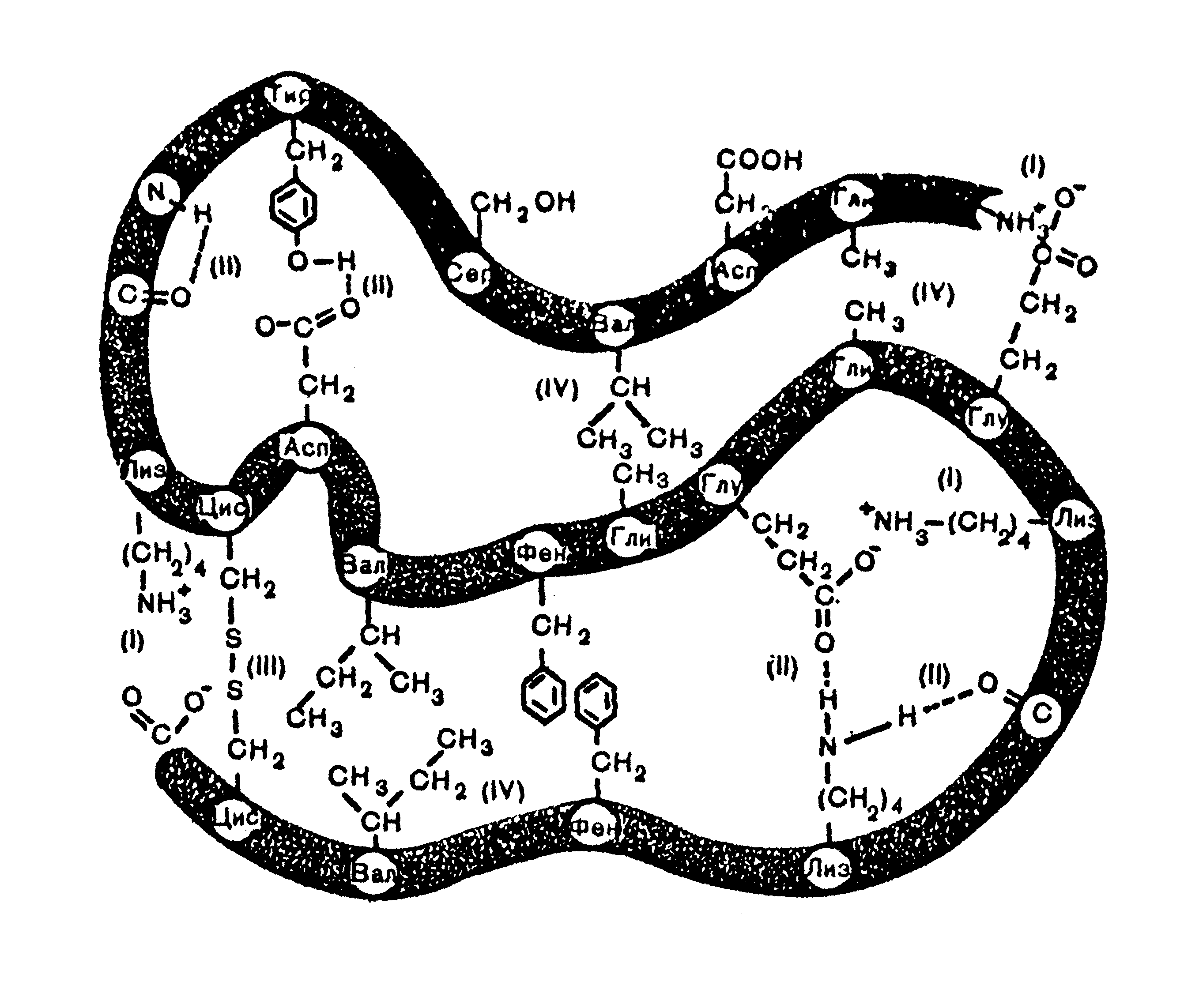
Мал. 6.12. Вплив взаємодій на просторове впорядкування білків.
Що стосується нуклеїнових кислот, то зараз досить багато відомо щодо третинної структури певних типів ДНК та РНК. На мал. 6.11 зображено просторове впорядкування ДНК в еукаріотичних клітинах. Видно, що основний елемент третинної структури утворений своєрідним “соленоїдом”, який складається з трьох великих витків, що мають діаметр приблизно 30 нм.
Характер взаємодій, які забезпечують просторове впорядкування білків, ілюструє мал. 6.12. Як видно з нього, стабілізація третинної просторової структури здійснюється як за рахунок ковалентних зв’язків та кулонівських (іон-іонних) взаємодій, так і за рахунок нековалентних зв’язків (водневих, дисперсійних, гідрофобних).
6.2.4. Четвертинна структура
Окремі поліпептидні ланцюги, що входять до складу білкової молекули та характеризуються певними первинною, вторинною і третинною структурами, можуть мати досить слабкі (нековалентні) зв’язки між собою.
Т
Мал.
6.13. Сферична білкова
макромолекула.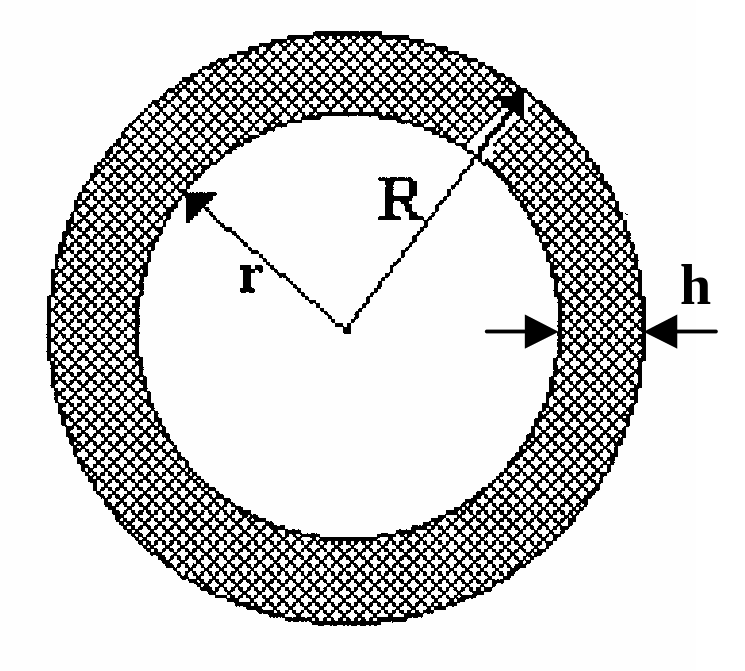
На закінчення цього параграфу розглянемо досить просту і водночас дуже корисну модель Фішера-Бреслера-Талмуда, яка дозволяє передбачити форму білкової макромолекули в залежності від відношення гідрофільних і гідрофобних груп, що входять до її складу.
Спочатку знайдемо те співвідношення, виконання якого гарантує сферичну глобулярну структуру білкової макромолекули. Нехай радіус сферичної глобули – R, радіус внутрішнього гідрофобного ядра – r, товщина гідрофільного шару – h (мал. 6.13). Обчислимо відношення об’ємів гідрофільної та гідрофобної частин в такій сферичній глобулі
![]() , (6.9)
, (6.9)
де враховані очевидні рівності для площі поверхні гідрофобного ядра S = 4r2, для об’єму гідрофільного шару завтовшки h на цьому ядрі Vгідрофільн = 4r2h, для об’єму самого сферичного гідрофільного ядра Vгідрофобн = 4r3/3, а також для радіусу всієї сферичної глобули R = r + h.
Звідси випливає, що для забезпечення сферичної форми білкової макромолекули повинно виконуватися наступне співвідношення для параметру bсф, що характеризує відношення об’ємів гідрофільної та гідрофобної частин:
![]() , (6.10)
, (6.10)
де
![]() – загальний об’єм сферичної
макромолекули, а коефіцієнт
– загальний об’єм сферичної
макромолекули, а коефіцієнт
![]() .
.
Таким чином, на кривій
залежності параметра bсф
від V1/3,
яка відповідає формулі (6.10), розташовуються
білкові макромолекули, що мають сферичну
форму (див. мал. 6.14). Якщо параметр
![]() набуває більшого
значення, ніж те, що визначається
формулою (6.10), тобто
набуває більшого
значення, ніж те, що визначається
формулою (6.10), тобто
![]() ,
(область над графіком bсф
= f
(V1/3)
на мал. 6.14), то білкова макромолекула
набуває не сферичної, а еліпсоїдальної
(фібрилярної) форми. Причина появи
фібрилярної структури полягає в тому,
що із зростанням кількості гідрофільних
амінокислотних залишків вони прагнуть
завдяки диполь-дипольним взаємодіям
покрити більшу площу, аніж площа
сферичної поверхні, яка є мінімальною
за величиною при фіксованому об’ємі.
,
(область над графіком bсф
= f
(V1/3)
на мал. 6.14), то білкова макромолекула
набуває не сферичної, а еліпсоїдальної
(фібрилярної) форми. Причина появи
фібрилярної структури полягає в тому,
що із зростанням кількості гідрофільних
амінокислотних залишків вони прагнуть
завдяки диполь-дипольним взаємодіям
покрити більшу площу, аніж площа
сферичної поверхні, яка є мінімальною
за величиною при фіксованому об’ємі.
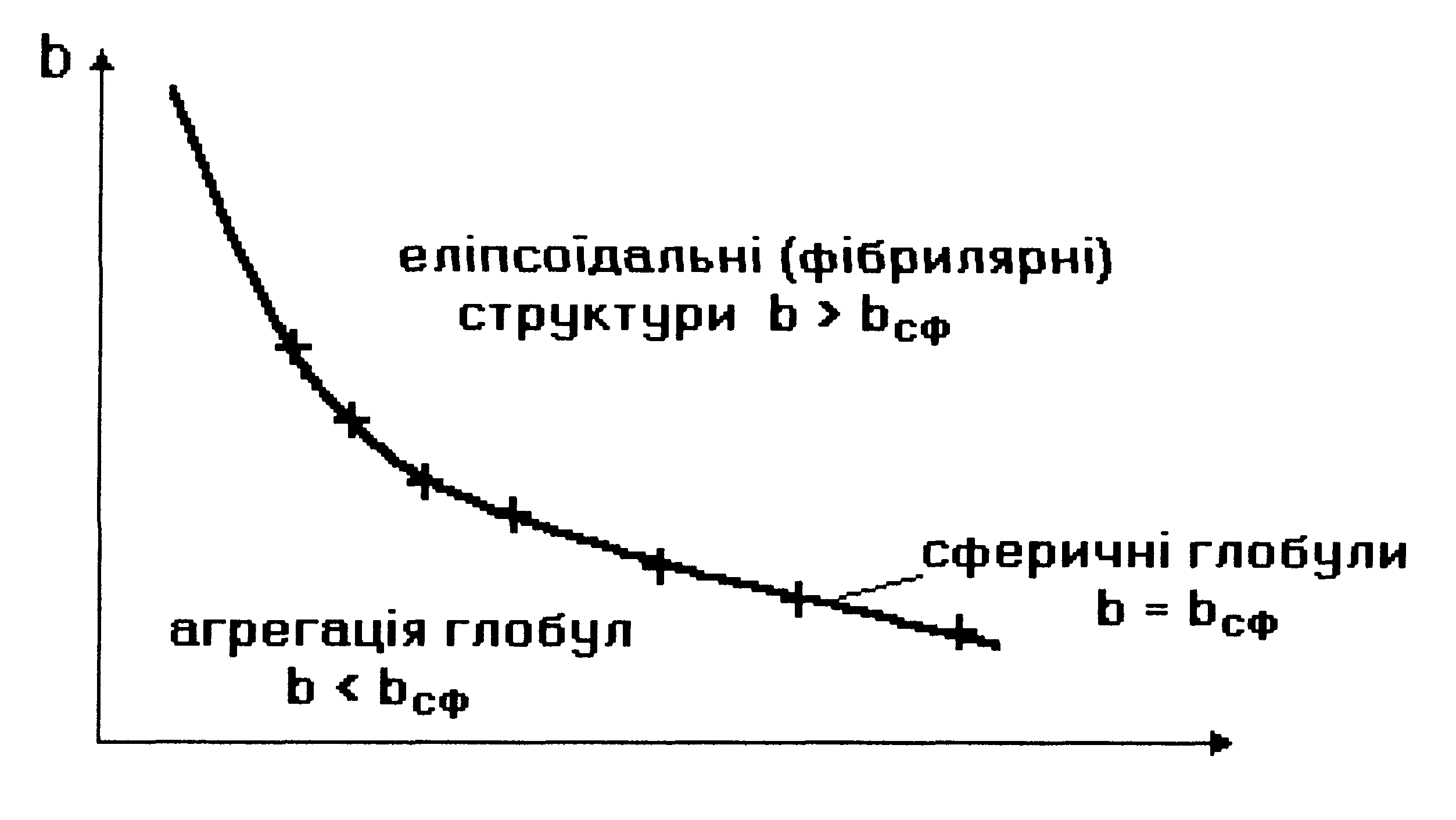
Мал. 6.14. Можливі форми білкових макромолекул в моделі Фішера-Бреслера-Талмуда.
В тому ж випадку, коли (область під кривою залежності bсф від V1/3 на мал. 6.14), кількості гідрофільних груп не вистачає навіть для покриття поверхні сферичної глобули. На сферичній поверхні залишаються гідрофобні області, які повинні “заховатися” від полярного оточення. Тому стає детермінованим процес (ймовірність якого прямує до одиниці) об’єднання глобул з утворенням четвертинної структури. Типовим проявом подібного механізму утворення четвертинної структури є молекула гемоглобіну, що складається, як вже зазначалося, з 4 глобул, саме завдяки гідрофобним взаємодіям.