

6.4.3. Активний транспорт
Поряд з пасивним транспортом у життєдіяльності клітини важливу роль відіграють процеси примусового переносу молекул та іонів з області малих концентрацій до області високих концентрацій, тобто активний транспорт. Завдяки активному транспорту підтримуються концентраційні градієнти, які необхідні для нормального функціонування клітини. Транспортні системи, які створюють необхідні концентраційні градієнти, називають насосами, або АТФ-азами. Відомі чотири основні системи активного транспорту:
1. Na+-К+-насос;
2. Са2+-насос;
3. Н+-насос;
4. Перенос протонів при роботі дихального ланцюга мітохондрій.
Активний транспорт, як і пасивний, забезпечується спеціальними структурами: каналами, переносниками, ферментами. При активному транспорті (на відміну від пасивного) вектор переміщення іонів співпадає за напрямком з вектором концентраційного градієнту, тобто з напрямком збільшення концентрації. Активний транспорт відбувається за рахунок енергії, що виділяється при гідролізі АТФ (комплексу Mg-АТФ2-) з утворенням молекул АДФ і неорганічного фосфату (Фн). Гідроліз АТФ здійснюється ферментом АТФ-азою. Джерелом молекул АТФ є процеси окислювального фосфорилювання, що відбуваються в мітохондріях.
Розглянемо основні етапи роботи Na+-K+ насосу (мал. 6.29). Як відомо, в нормально функціонуючій клітині концентрація іонів К+ перевищує їх концентрацію в міжклітинному середовищі, а для іонів Na+ це співвідношення обернене. Наприклад, у нервовому волокні кальмара:
[К+]е = 0.010 моль/л, [К+]i = 0.340 моль/л,
[Na+]е = 0.463 моль/л, [Na+]i = 0.049 моль/л,
тобто [К+]i > [К+]е , [Na+]i < [Na+]е .
N a+-K+-ATФ-aзa
локалізована у зовнішніх плазматичних
мембранах клітин. Через те, що
Na+-K+-ATФ-aзa є гідрофобним
білком, вона створює комплекси з ліпідами
бішару. Саме тому активність інтегрального
білка Na+-K+-ATФ-aзи залежить
від фізико-хімічного стану мембрани.
Встановлено, що для
функціонування Са2+-АТФ-ази
важливо, щоб ліпідне оточення
було рідким, на відміну від Na+-K+-ATФ-ази,
для якої важлива впорядкованість
бішару.
a+-K+-ATФ-aзa
локалізована у зовнішніх плазматичних
мембранах клітин. Через те, що
Na+-K+-ATФ-aзa є гідрофобним
білком, вона створює комплекси з ліпідами
бішару. Саме тому активність інтегрального
білка Na+-K+-ATФ-aзи залежить
від фізико-хімічного стану мембрани.
Встановлено, що для
функціонування Са2+-АТФ-ази
важливо, щоб ліпідне оточення
було рідким, на відміну від Na+-K+-ATФ-ази,
для якої важлива впорядкованість
бішару.
Перенос двох іонів К+ всередину клітини і трьох іонів Na+ зовні приводить до переносу одного позитивного заряду із цитоплазми в оточуюче середовище. Внаслідок цього всередині клітини виникає негативний електричний потенціал. Тому Na+-K+-насос називають електрогенним.
|
Мал. 6.30. Співвідношення напрямків потоків іонів Na+ та К+ та відповідних градієнтів концентрацій, а також градієнти електричного потенціалу при активному транспорті. |
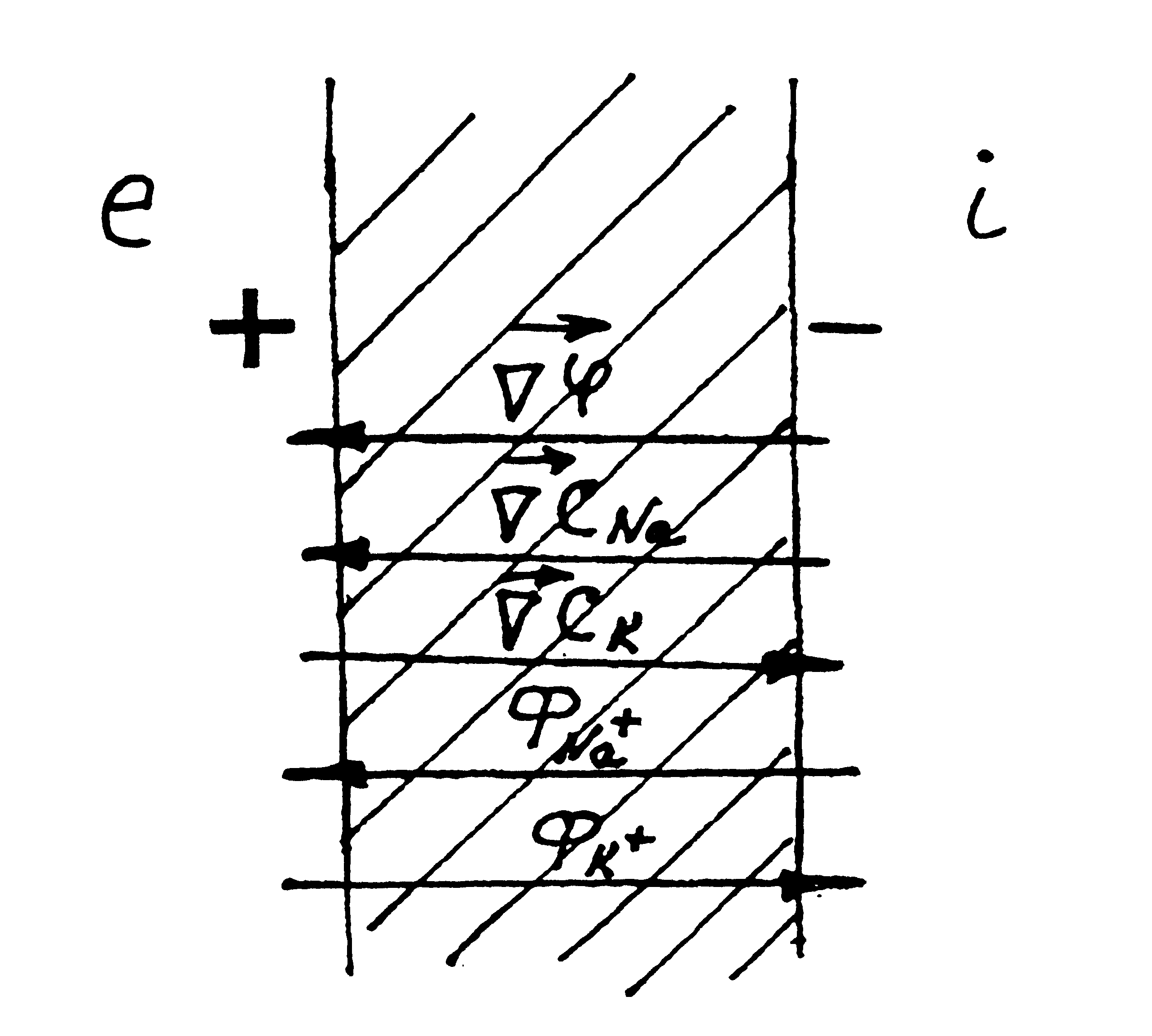
Таким чином, при активному транспорті
потік іонів Na+ (ФNa)
співпадає з напрямками градієнта
концентрації Na+ і градієнта
електричного потенціалу
![]() ,
а потік іонів К+ (ФK) –
напрямлений по градієнту концентрації
К+, але проти
(мал. 6.30).
,
а потік іонів К+ (ФK) –
напрямлений по градієнту концентрації
К+, але проти
(мал. 6.30).
Біологічні потенціали
Генерація і поширення електричних потенціалів – найважливіше явище в живих клітинах і тканинах, яке лежить в основі збудження клітин, регуляції внутрішньоклітинних процесів, м’язового скорочення, роботи нервової системи. Порушення електричних характеристик окремих клітин, нервових волокон, тканин приводить до захворювання.
Різниця потенціалів між водними фазами по обидва боки мембрани (або мембранний потенціал) виникає внаслідок процесів переносу, тобто процесів пасивного і активного транспорту.
Потенціал спокою – різниця потенціалів між цитоплазмою і навколишнім середовищем у нормально функціонуючій незбудженій клітині, що обумовлена відміною у концентраціях іонів по обидва боки мембрани.
Потенціал дії – різниця потенціалів, що виникає при збудженні клітини і обумовлена зміною проникності мембрани для іонів.
Загальна картина розподілу потенціалів на межах мембрани з водними фазами (внутрішньоклітинній і позаклітинній) подана на мал. 6.31. З цього рисунка випливає існування чотирьох характерних різниць потенціалів:
Мал. 6.31. Зміна
електричного потенціалу
при наближенні до мембрани і
в самій мембрані.
s2 – міжфазний стрибок потенціалу на внутрішній границі мембрани;
– різниця потенціалів на границях мембрани;
м = (e – i) – мембранний потенціал, або різниця потенціалів, між водними фазами по обидва боки мембрани. При цьому
![]() .
.
Для симетричної мембрани
![]() ,
,
тоді
![]() ,
,
тобто
![]() .
.
Міжфазні стрибки потенціалу (s1, s2) обумовлені поверхневим зарядом, що його створюють “голівки” фосфоліпідів, та здатністю мембрани зв’язувати іони з навколишньої водної фази. Спадає потенціал за законом:
![]() ,
,
де – радіус Дебая, що дорівнює тій відстані від границі мембрани, на якій потенціал зменшується в е разів. Радіус Дебая залежить від концентрації іонів у водній фазі і складає приблизно 0.8 нм, тобто 10% від товщини мембрани.
Мембрана за своїми електричними властивостями подібна до конденсатора. Потенціал змінюється всередині мембрани приблизно за лінійним законом, тобто напруженість поля є майже сталою величиною:
![]() .
.
Походження мембранних потенціалів можна пояснити, виходячи з особливостей розподілу і дифузії іонів з урахуванням їх проникності крізь мембрану.