70. Биосинтез белка в дрожжевой клетке.
C 2H12O6 2CH3COCOOH
2H12O6 2CH3COCOOH
гликолиз пируват пируват-декар
боксилаза
2 CH3CH2OH 2CH3CHO
Алкаголь-дегидрогеназа
-
Спиртовое брожение. Преоблад. продукт - этиловый спирт. Возбудители: дрожжи Saccharomyces ccrevisiac, S.uvarum, Schizosacci aromyces pombe, бактерии Zymomonas mob His. Субстраты - глюкоза, фруктоза, сахароза, мальтоза, галактоза. Изв-но 3 сп-ба спиртового брож-я, отлич-ся др. от др. ем, какой из 3-х катабол. путей расщепления глюкозы реализ-ся - гликолиза (дрожжи), пентозофосф. пути (гетерофермент. молочнокислые бактерии, прод-ты брож-я - этанол и молочная к-та) или путь Энтнера-Дудорова (бактерии Zymomonas mobilis, прод-ты брож-я -этанол, ССЬ, небольшое кол-во молочной к-ты). Чаще клетки дрожжей исп-т гликолиз, и его конечный продукт (пируват) служит субстратом спиртового брож-я. В ходе 2-х р-ций брожения пируват превр-ся в этанол. Проц-с сопровожд-ся регенерацией 2-х мол-л НАД4, к-рые снова могут уч-ть в окислении глюкозы до пирувата.
Следуе отметить, что_АТФ запас-ся только в гликолизе, в р-циях собственно спирт брож-я этого не происх-т. Этанол удал-ся из клеток. м-ду кол-вом этанола и СО2 собл-ся эквивал-ть - на 2 мол-лы этанола образ-ся 2 мол-лы СО2. Кроме этанола образ-ся примеси - глицерол, уксусны альдегид, др. альдег, эфиры, кетоны, спирты, орг. к-ты.
72. Биохимия образования этанола дрожжами.
В связи с тем, что дрожжи ассимилируют готовые органические соединения и не усваивают Н2СО3, они относятся к гетеротрофным микроорганизмам, а по типу дыхания − к факультативным анаэробам, т. е. способны развиваться как в анаэробных условиях в отсутствии кислорода, так и в аэробных условиях.
При спиртовом брожении происходят ферментативные превращения гексозных моносахаридов в анаэробных условиях, приводящие к их неполному окислению и сопровождающиеся выделением энергии. В этих окислительных реакциях акцептором водорода является не кислород, а промежуточные продукты превращений углеводов, например уксусный альдегид. Биохимические процессы анаэробиоза протекают внутри дрожжевой клетки, куда через полунепроницаемую оболочку поступают моносахариды и необходимые неорганические питательные вещества. Продукты метаболизма − этанол, диоксид углерода и примеси выделяются из клетки в сбраживаемое сусло.
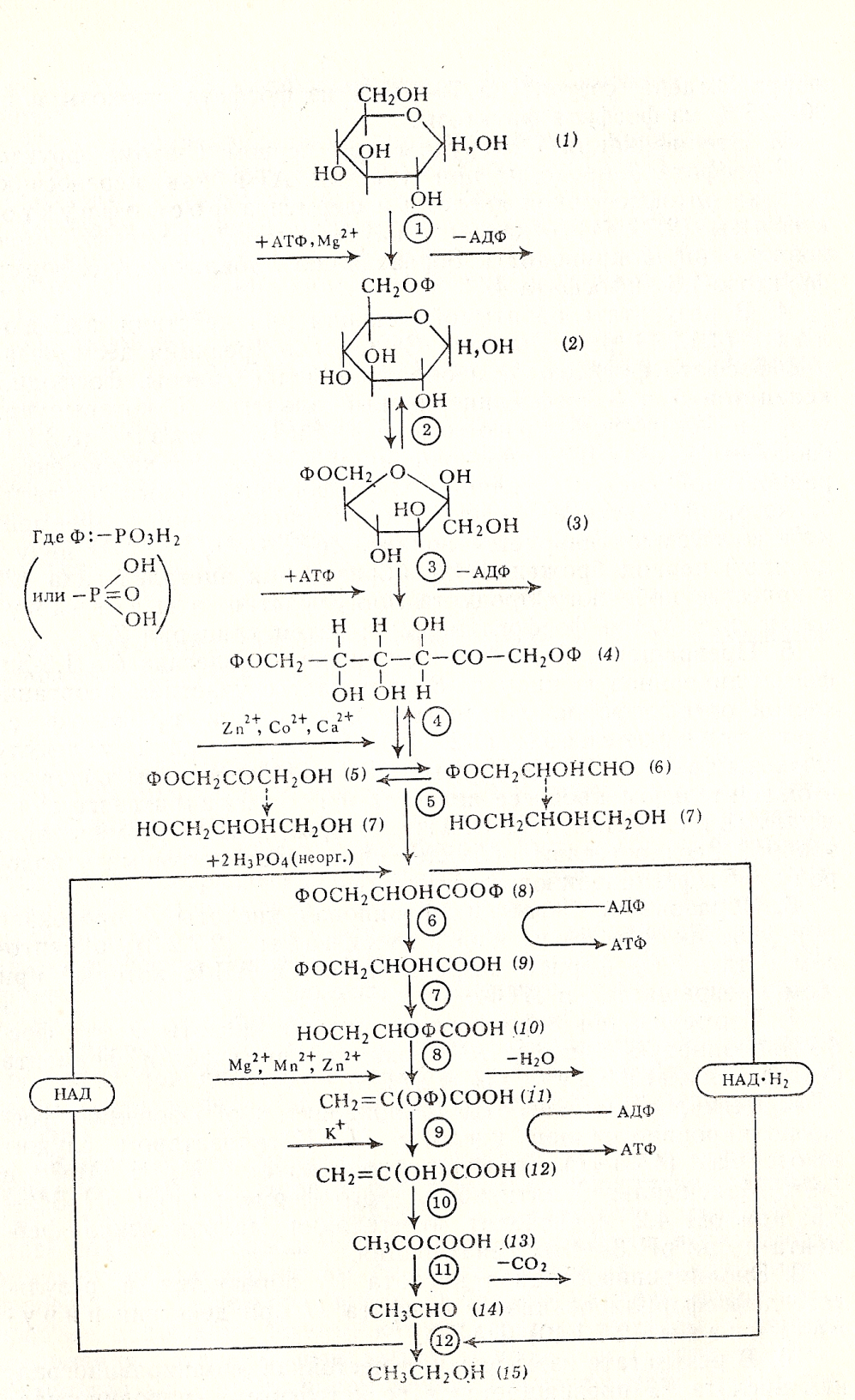
Основные стадии химизма спиртового брожения на примере превращений D-глюкозы, которая является основным углеводным компонентом гидролизных субстратов.
Фосфорилирование D-глюкозы 1 происходит при участии в качестве катализатора фермента гексокиназы. Каталитическая активность этого фермента повышается в присутствии Mg2+. Переносчиком группы − РО3Н2 является трифосфат адениновой кислоты (аденозинтрифосфат − АТФ), который превращается в аденозиндифосфат (АДФ). D-Глюкоза в этой реакции этерифицируется в пиранозной форме; ее реакционная способность при этом возрастает. Скорость образования фосфорного эфира D-глюкозы (глюкозо-6-фосфата) 2 определяет общую скорость брожения.
Изомеризация глюкозо-6-фосфата 2 в фруктозо-6-фосфат 3 протекает при участии фермента глюкозофосфатизомеразы. Равновесная система, получившая название эфира Эмдена, состоит на 70−75% из фосфата глюкозы и на 30−25 % из фосфата фруктозы.
Этерификация ОН-группы при первом С-атоме фруктозо-6-фосфата 3 проходит при участии АТФ как переносчика остатка ортофосфорной кислоты и фермента фосфофруктокиназы в качестве катализатора. Раскрытие пиранозного цикла приводит к образованию лабильной оксоформы фруктозо-1,6-дифосфата 4.
4.В результате обратимой реакции при действии альдолазы расщепляется С−С-связь (реакция десмолиза) в дифосфате фруктозы 4 и образуются две триозы: фосфодиоксиацетон 5 и 3-фосфоглицериновый альдегид 6, которые при участии фермента триозофосфатизомеразы способны к взаимным превращениям. Основным компонентом равновесной системы является 3-фосфоглицериновый альдегид 6, который участвует в дальнейших превращениях. Фермент альдолаза активируется ионами Zn2+, Со2+, Са2+. В индукционный период брожения (до образования ацетальдегида 12) в качестве побочного продукта при участии альдегидмутазы образуется фосфоглицерин и затем глицерин 7.
Превращение 3-фосфоглицеринового альдегида 6 в 1,3-дифос-фоглицериновую кислоту 8 протекает с участием неорганической ортофосфорной кислоты в присутствии триозофосфатдегидрогеназы. В расчете на 1 молекулу глюкозы требуется 2 молекулы Н3Р04. Переносчиком водорода в окислительном процессе является никотинамидаденин-иуклеотид (НАД), который как кофермент анаэробной дегидрогеназы обладает способностью отнимать водород от фосфоглицеринового альдегида.
Образование 3-фосфоглицериновой кислоты 9 протекает при участии фосфоглицераткиназы; акцептором остатка фосфорной кислоты является АДФ, который при этом превращается в АТФ.
Изомеризация 3-фосфоглицериновой кислоты 9 в 2-фосфоглицериновую кислоту 10 проходит при участии фермента фосфоглицеромутазы.
Дегидратация кислоты 10 приводит к образованию фос-фоэнолпнровиноградной кислоты 11. Катализатором служит энолаза, активируемая ионами Mg2+, Мп2+ и Zn2+. Максимальная активность этого фермента при рН 5,2− 5,5; при рН 4,2 происходит агрегатация макромолекул фермента и при рН 3−4 их денатурация.
Энолпировиноградная кислота 12 образуется в результате дефосфорилирования ее фосфата 11 при действии пируваткиназы и АДФ.
Декарбоксилирование пировиноградной кислоты 13 при участии пируватдекарбоксилазы приводит к образованию уксусного альдегида 14.
Восстановление ацетальдегида 14 в этанол 15 протекает при участии НАД-2Н и фермента алкогольдегидрогеназы. При этом кофермент регенерируется путем окисления в НАД.
По такой же схеме происходит биосинтез этанола и из других гексоз гидролизного субстрата − D-маннозы и D-галактозы.
Из рассмотренной схемы видно, что для нормального протекания спиртового брожения необходимо наличие в субстрате неорганического фосфорного питания и макро- и микроэлементов К, Mg, Са, Mn, Zn и Со, активирующих ферменты.
Суммарное стехиометрическое уравнение спиртового брожения, установленное еще Гей-Люссаком,
С6Н1206 − > 2С2Н5ОН + 2С02
может быть представлено с учетом роли АТФ в жизненных процесса следующим образом:
С6Н1206 + 2АДФ + 2Н3Р04 −> 2С2Н5ОН + 2С02 + 2АТФ
Изменение свободной энергии в этом процессе составляет более 200 кДж/моль, в том числе 83,4 кДж/моль улавливается в АТФ и используется для осуществления жизненных функций микроорганизмов. Остатки фосфорной кислоты в АТФ присоединены макроэргическими связями, при расщеплении которых освобождается примерно в 3 раза больше энергии, чем при расщеплении обычных химических связей. Освобождение энергии макроэргических связей происходит при переносе − РО3Н2 групп, а не путем гидролиза, при котором свободная энергия переходит в тепловую.
В анаэробных условиях происходит медленное размножение дрожжей. При биосинтезе белка и других компонентов клеток кроме энергии затрачивается часть питательных веществ субстрата.
При спиртовом брожении в качестве примеси образуется глицерин, выход которого возрастает в присутствии бисульфита (например, NaHS03, KHS03) или в щелочной среде. В составе спиртовой бражки содержится ацетальдегид, являющийся промежуточным продуктом биосинтеза этанола. Среди многочисленных легколетучих примесей бражки найдены альдегиды, простые и сложные эфиры, кетоны, спирты, кислоты и другие соединения. Значительный выход эфиров при спиртовом брожении связан с протеканием реакций этерификации, катализируемых ферментами эстеразами. Основной компонент эфирной фракции − этилацетат 17, образуется при взаимодействии этанола 15 и уксусной кислоты 16.
С2Н5ОН + СН3СООН <=> CH3CO2C2H5 + Н20
15 16 17
В процессе обмена веществ, главным образом при размножении микроорганизмов, в качестве побочных продуктов брожения образуются высшие спирты. Их выход возрастает при наличии аминокислот, которые попадают в субстрат из исходного сырья, а также при автолизе отмерших дрожжевых клеток. В условиях брожения происходит декарбоксилирование и гидролитическое дезаминирование аминокислот с образованием пропанола, изобутанола, амилола, изоамилола и других спиртов, которые при ректификации образуют фракцию сивушных масел.
Основным компонентом сивушных масел является изоамиловый спирт 21, который образуется из лейцина 18:
( (СН3)2СНСН2СН (NH2) СООН + СНзСОСООН −> СН3СН (СН3) СН2СОСООН + 18 13 19
+ CH3CH(NH2)COOH
20
-С02 +Н2
СОН3СН(СН3)СН2СОСОО −−> (СН3)2СНСН2СНО −−> СН3СН(СН3)СН2ОН
19 20 21
На первой стадии происходит процесс переаминирования − обмен аминогруппой между аминокислотой 18 и кетокислотой (пировиноградной кислотой 13, при этом образуется аланин 20 и изопропилвиноградная кислота 19, декарбоксилирование которой приводит к изовалериановому альдегиду 20, при восстановлении которого образуется изоамилол 21. Все стадии процесса протекают при участии ферментов.