

Смертность.
Этот показатель характеризует гибель особей популяций за определенный период времени. Так же, как и в случае с рождаемостью, смертность можно выразить числом особей, погибших за определенный период.
Различают:
специфическую смертность - число смертей по отношению к числу особей, составляющих популяцию.
экологическую или реализуемую, смертность - гибель особей в конкретных условиях среды (величина непостоянная, изменяется в зависимости от состояния природной среды и состояния популяции).
Смертность выражают числом особей, погибших за определенное время, выраженным в процентах от начальной численности популяции.
![]()
Как и в случае с рождаемостью, необходимо принять минимальную смертность за теоретическую константу.
Существует некая минимальная величина, характеризующая гибель особей в идеальных условиях, когда на популяцию не воздействуют лимитирующие факторы. В этих условиях максимальная продолжительность жизни особей равна их физиологической жизни, которая в среднем выше экологической продолжительности жизни.
Даже в идеальных условиях в любой популяции происходит смерть от старости, то есть имеет место какая-то минимальная смертность, определяемая физиологической продолжительностью жизни особей.
В природе средняя продолжительность жизни особей намного меньше, чем потенциальная, и фактическая величина смертности выше минимальной.
Смертность, как и рождаемость, особенно у высших организмов, сильно варьируется с возрастом, поэтому при изучении смертности организмов популяцию целесообразно разделить на возрастные группы.
Различают следующие типы таблиц выживания:
1.Когортные (динамические). Прослеживается история существования только одной когорты особей (от момента появления до гибели всех ее особей). При этом когортой мы называем группу особей родившихся в течение определенного промежутка времени. Часто применяется для изучения популяций однолетних организмов (насекомых).
2.Статические таблицы выживания. Вместо наблюдения за отдельной когортой ведут наблюдение за возрастной группой. При этом расчитывается смертность во всех возрастных группах.
Сложность построения таких таблиц заключается в невозможности реконструкции численности особей разных возрастных групп, с момента их рождения.
Они могут содержать следующие графы:
возраст (временные промежутки, стадии развития (для личинок));
число особей доживших до начала возрастного интервала, nx;
доля особей доживших до возраста x, lx,;
число особей, погибших в каждом возрастном интервале (x) – (x+1), dx;
смертность в течение интервала x, qx;
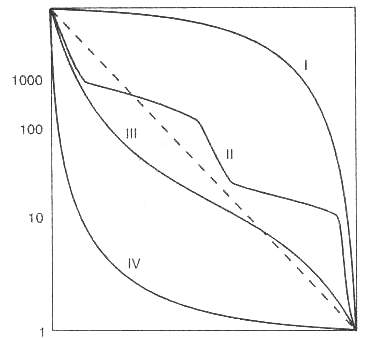
Если построить график зависимости lx от Х, то получим
кривую выживания (по числу доживших до данного возраста организмов популяции).

Выделяют три типа кривых выживания:
сильновыпуклые кривые (I) - характерны для видов организмов в популяциях, смертность которых почти до конца жизненного цикла остается низкой. Такой тип кривых выживания характерен для многих видов крупных животных, в том числе и для человека;
сильновогнутые кривые (IV) - характерны для видов организмов в популяциях, смертность которых чрезвычайно высока на ранних стадиях развития, а в дальнейшем становится величиной постоянной, но достаточно высокой. Например, 1-й личиночный возраст у насекомых, при этом особи погибают в больших количествах, а гибель в последующих возрастных группах снижается.
кривые промежуточного типа (II-III) относятся к тем видам, у которых смертность для каждой возрастной группы - величина постоянная.
|
По оси абцисс (горизонт.) - Возраст, в %% от продолжительности жизни По оси ординат (верт.) – число выживших на 1000 особей (или % выживших от исходной популяции). |
|
|
8. Экспоненциальная модель популяционного роста. Биотический потенциал. Зависимость скорость экспоненциального роста от характеристик организма (размера и др.), обеспеченности ресурсами и условий среды.
Экспонциальная модель популяционного роста. dN/dt = rN. В такой форме записи подчеркивается, что скорость роста численности популяции пропорциональна самой численности. Если r = const, то рост происходит по экспоненциальному закону. В том случае, когда величины численности приводятся в логарифмическом масштабе, график экспоненциального роста приобретает вид прямой линии. Поэтому иногда экспоненциальный рост называют логарифмическим. Уравнение экспоненциального роста в логарифмической форме имеет следующий вид: lnNt = lnN0 + rt, т. е., по сути, это просто уравнение прямой, причем коэффициент r характеризует угол наклона ее к осям.
Во многих популярных руководствах по экологии говорится, что экспоненциальный рост популяций возможен только в особо оптимальных условиях, при отсутствии каких-либо ограничивающих факторов. Это не совсем верно, поскольку, как уже подчеркивалось выше, единственное (необходимое достаточное) условие такого роста — постоянство коэффициента г, отражающего, по сути дела, скорость размножения данных организмов. Так, например, проведя серию наблюдений за ростом популяций какого-либо одноклеточного организма в разных температурных условиях, нетрудно заметить, что с уменьшением температуры скорость деления клеток падает, но экспоненциальный характер роста сохраняется во всех вариантах.
Скорость экспоненциального роста популяций может использоваться как показатель благоприятности условий среды для данных организмов. Очевидно, например, что с повышением температуры у пойкилотермных животных эта скорость растет, но с приближением к границе толерантности снижается. У разных видов характер этих изменений может быть различным.
Математические формулы, описывающие закономерности экспоненциального роста, были приведены А. Лоткой в 1920-х гг., но основной принцип экспоненциального роста (или, как его чаще называли, роста в геометрической прогрессии) был известен уже очень давно. О том, что принципиально такой рост возможен, упоминали Ж. Бюффон и К. Линней. До них, еще в конце XVII в. особенности экспоненциального роста были очевидны одному из основателей демографии — Дж. Гранту. О геометрической прогрессии как о некотором законе роста народонаселения писал и Томас Мальтус (1766—1834), труды которого оказали большое влияние на формирование взглядов двух исследователей, выдвинувших идею естественного отбора,— Ч. Дарвина и А. Уоллеса. Именно врожденная способность любой группы организмов неограниченно увеличивать свою численность по экспоненциальному закону служит одной из основных предпосылок теории естественного отбора.
Биотический потенциал в экологии, способность вида противостоять неблагоприятным воздействиям внешней среды. Термин введён американским экологом Р. Чепменом (1925) в связи с проблемой динамики численности животных. По Чепмену, Биотический потенциал - количественное выражение способности организмов противостоять сопротивлению внешней среды. Согласно его теории, потенциальная плодовитость животных не реализуется, поскольку она подавляется односторонним воздействием внешней среды, с которой организмы находятся в антагонистических отношениях. По современным воззрениям, такая точка зрения выглядит упрощённой. Изменения плодовитости и выживания животных происходят как под влиянием абиотических факторов, так и в результате межвидовых и внутривидовых взаимоотношений. Большую роль в этих процессах играют внутрипопуляционные механизмы, обеспечивающие активную реакцию популяции на внешние воздействия.
.
9. Колебания численности популяции. Механизмы регуляция численности популяции. Типы динамики популяций. Стратегии развития популяций. K и r – стратегии. Экостратегии по Л. Г. Раменскому.
По достижении заключительной фазы роста размеры популяции продолжают колебаться от поколения к поколению вокруг некоторой более или менее постоянной величины. При этом численность одних видов изменяется нерегулярно с большой амплитудой колебаний (насекомыевредители, сорняки), колебания численности других (например, мелких млекопитающих) имеют относительно постоянный период, а в популяциях третьих видов численность колеблется от года к году незначительно (долгоживущие крупные позвоночные и древесные растения).В природе в основном встречаются три вида кривых изменения численности популяции: относительно стабильный, скачкообразный и циклический. Виды, у которых численность из года в год находится на уровне поддерживающей емкости среды, имеют достаточно стабильные популяции. Такое постоянство характерно для многих видов дикой природы и встречается, например, в нетронутых тропических влажных лесах, где среднегодовое количество осадков и температура изменяются день ото дня и из года в год крайне мало.У других видов колебания численности популяций носят правильный циклический характер. Ряд видов, таких, как енот, в основном имеют достаточно стабильные популяции, однако время от времени их численность резко возрастает (подскакивает) до наивысшего значения, а затем резко падает до некоторого низкого, но относительно стабильного уровня. Эти виды относят к популяциям со скачкообразным ростом численности Внезапное увеличение численности происходит при временном повышении емкости среды для данной популяции и может быть связано с улучшением климатических условий (факторов) и питания или резким уменьшением численности хищников (включая охотников). После превышения новой, более высокой емкости среды в популяции возрастает смертность и ее размеры резко сокращаются.
Численность каждого отдельного вида организмов сохраняет равновесное положение в своей среде обитания. Одновременно в течение сезона численность популяции в разные годы колеблется в ту или иную сторону от этого равновесного, сравнительно неизменного среднего значения. Такое равновесие за какой-то период времени сохраняется из-за действия на популяцию всех факторов внешней среды (естественное регулирование).Наблюдаемые в природе непрерывные изменения численности организмов являются результатом взаимодействия двух процессов: модификации и регуляции (Г. А. Викторов, 1976).Модификация обусловлена действием на популяцию абиотических факторов среды (ветра, влажности воздуха и др.). Однако абиотические факторы не могут реагировать на изменение плотности популяции организмов в сторону усиления или ослабления своего стрессового воздействия, т. е. менять его по принципу обратной связи. Так, при низких зимних температурах может наблюдаться высокая смертность эмбрионов в яйцах яблонной медяницы, зимующих на ветвях яблони. Однако независимо от смертности насекомого температура воздуха не повышается, и, если погодные условия не изменяются, эмбрионы погибают.Существуют две группы регулирующих механизмов — внутривидовые и биоценотические.Среди внутривидовых регуляторных механизмов широко известно угнетающее влияние друг на друга особей одной популяции, использующих одни и те же ограниченные пищевые ресурсы. Иногда этот процесс осложняется каннибализмом и другими формами агрессивного поведения.Более сложны внутривидовые регуляторные механизмы, основанные на сигнальном действии роста плотности популяции. Это происходит до истощения пищевых ресурсов, предотвращающих конкуренцию за них.
Типы динамики популяций. Механизмы динамики популяций
Типы динамики популяций:
Стабильный – небольшая амплитуда изменения численности; характерен для крупных животных с продолжительностью жизни 15-20 и более лет.
Лавильный тип – колебания численности могут достигать нескольких десятков раз, для видов с продолжительностью жизни 5-10 лет.
Эфемерный тип – изменения численности в сотни раз, для видов с продолжительностью жизни 1 год и менее.
Связь типа динамики популяций и биологических особенностей организмов лежит в основе концепции экостратегий (Макартор, 1967). Выделены следующие типы стратегии:
r-стратеги – отбор на количество: максимальная скорость в момент освоения территории; имеют явное преимущество при заселении территории;
k-стратеги – отбор на качество: выигрывают конкуренцию в зрелых сообществах.
Л. Г. Раменский выделял три типа экостратегий: эксплеренты, виоленты и патиенты.
Существуют три основных концепции объяснения изменения численности популяций:
Регуляционизм – каждая популяция обладает своим уровнем численности, отклонения с ту или иную сторону регулируются биотическими факторами;
Стохастизм – стабильно многолетнее среднее значение численности популяции (тем более постоянно, чем больше период), поддерживаемое абиотическими факторами регуляции;
Саморегуляция.
Механизмы динамики популяций:
Случайные явления;
Климатическая теория;
Трофическая теория (из-за изменения пищи);
Популяционная теория.
10. Внутривидовая конкуренция. Эксплуатация и интерференция. Зависимость смертности и плодовитости от плотности популяции. Асимметричность конкуренции. Логистическая модель роста популяции.
Конкуренция
Введем рабочее понятие конкуренции – как взаимодействия между особями которое вызвано сходными потребностями в ограниченном ресурсе, что приводит к снижению выживаемости, скорости роста (или размножения) конкурирующих особей.
Внутривидовая конкуренция (ВВК)
Внутривидовая конкуренция - это соперничество особей одного вида за ресурс, когда его не хватает. В качестве примера такой конкуренции рассмотрим гипотетическое сообщество - процветающую популяцию кузнечиков одного вида. Разыскивая и потребляя пищу, кузнечики расходуют энергию и подвергают себя риску быть съеденными. При увеличении плотности популяции для обеспечения жизнедеятельности организмов потребуются большие затраты времени и энергии. В этом случае возрастает внутривидовая конкуренция, что приводит к повышению энергетических затрат и к снижению скорости потребления пищи. В результате шансы выжить уменьшаются.
Аналогична ситуация и у растений. Один проросток растения с большей вероятностью доживет до репродуктивной зрелости, чем проросток, окруженный густой порослью. Даже если он не погибнет, то почти наверняка будет мелким, неразвитым, даст мало потомства. Следовательно, повышение плотности популяции уменьшает вклад каждой особи в следующее поколение.
Признаки внутривидовой конкуренции:
ВВК приводит к снижению скорости потребления ресурса в расчете на 1-у особь, что уменьшает ее плодовитость в конечном счете.
При ВВК особи могут непосредственно не взаимодействовать друг с другом, а реагируют лишь на уровень ресурса оставленного конкурирующими особями. Такой тип конкуренции называют эксплуатационной.
При непосредственном взаимодействии особей друг с другом имеет место так называемая интерференционная конкуренция. Например среди подвижных территориальных животных (охрана территории).
Неравноценность конкурирующих особей (хотя казалось бы они должны быть равноценны). Они во многом сходны по основным характеристикам и особенностям использования среды, но результаты взаимодействия между конкурирующими особями неодинаковы. При этом при ВВК наблюдается асимметричность (более сильный экземпляр будет затенять слабый). При этом, хотя конечным результатом конкуренции и является уменьшение числа потомков, но это не означает снижение индивидуальной приспособленности (т.е. относительного вклада), особенно для самых сильных конкурентов. Следовательно ВВК не может «вредно влиять» на всех особей данного вида.
Возможное влияние на особь при ВВК тем сильнее, чем большее число конкурентов взаимодействует. Поэтому результаты ВВК следует считать зависимыми от плотности.
При ВВК смертность и плодовитость зависят от плотности популяции.
На рис. 1 (а,б,в) паказаны результаты зависимости смертности от плотности популяции мучного хрущака.
На участке (1) при низком значении плотности популяции результаты внутривидовой конкуренции отсутствуют.
На участке (2) видны результаты внутривидовой конкуренции, при этом, смертность растет, а число выживших почти не меняется.
На участке (3) наблюдается жесткая внутривидовая конкуренция, так как растет и удельная смертность и число погибших особей и падает чило выживших особей.
Показатель r - фактическая разность между специфической мгновенной скоростью рождений (b) и специфической мгновенной скоростью гибели организмов (d):
r = b - d
b= dNm / dt - мнгновенная скорость рождаемости d=dNn/ dt – мнгновенная скорость смертности
Скорость изменения численности (r) изменяется линейно и при достижении определенной численности популяции становится = 0.

Существует несколько способов регуляции численности, через зависимые от плотности факторы, при этом данные факторы, как мы видели меняются нелинейно:




При сравнении различных популяций, равенство их численностей, даже равновесных (то есть К) ничего не говорит о природе этого равновесия без учета динамики их рождаемости и смертности.
Друггая модель описывающая ВВК, показана на рисунке 2:

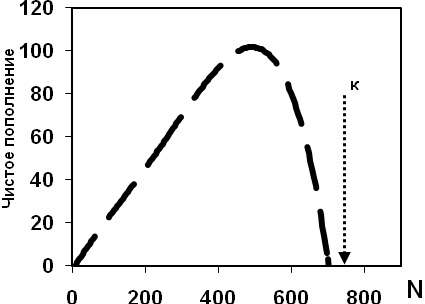
Чистая скорость пополнения – число особей на которое изменяется численность популяции за определенный промежуток времени.
В целом, пополнение популяции невелико кагда размножаются лишь некоторые особи и когда наблюдается интенсивная ВВК. При этом скорость пополнения максимальна лишь в определенный промежуток плотности.
Чистое пополнение всегда имеет вид колоколообразой кривой, форма которой является видоспецифичной.
Таким образом, ВВК имеет тенденцию регулировать численность популяции, при этом числая скорость роста популяции максимальна при промежуточных значениях плотности. В результате кривая популяционного роста имеет S-образный тип.
Асимметричная конкуренция.
ВВК может существенно влиять на различия особей внутри популяции. мы имеем дело не со средней особью, а с индивидуальностями.
Например, распределение массы растений льна относительно средней величины в зависимости от плотности посева.
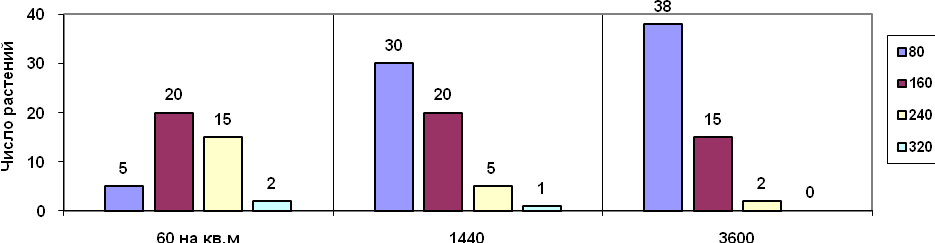
Однако асимметричная конкуренция может приводить к востановлению симметрии распределения в результате гибели слабых особей (примеры с «многоугольниками Тиссена» при прорастании семян и бокоплавах). При этом размер индивидуальных участков увеличивается и распределение выравнивается.
Территориальность является одной из форм асимметричной видовой конкуренции.
Размеры индивидуального участка зависят от соотношения затрат на поиск пищи на этом участке и выигрыша (количество пищевого русурса - нектар у колибри). Чем выше концентрация ресурса, тем меньше индивидуальный участок.
Модель предложена Мальтусом в 1798 г. в его классическом труде "О законе роста народонаселения".
Томас Роберт Мальтус (1766-1834) - известный английский демограф и экономист, обратил внимание на тот факт, что численность популяции растет по экспоненте (в геометрической прогрессии), в то время как производствопитания растет со временем линейно (в арифметической прогрессии), из чего сделал справедливый вывод, что рано или поздно экспонента обязательно "обгонит" линейную функцию, и наступит голод.
Если среда не лимитирует популяцию, то динамика численности популяции описывается дифференциальным уравнением: dN/dt = rN (скорость роста численности пропорциональна числу особей), которое имеет решение в виде: N = N0ert.
Здесь N0 - численность в начальный момент времени, t - время, r - коэффициент роста популяции, определяемый как разность между удельной рождаемостью b (в расчете на одну особь) и удельной смертностью d в данный момент времени, то есть r = b - d.
Максимально возможное значение коэффициента роста популяции rmax, достигаемое при наиболее благоприятных условиях среды, называется биотическим потенциалом популяции.
Обычно биотический потенциал тем выше, чем ниже уровень организации организмов. Так дрожжевые клетки, размножаясь делением, при условии реализации биотического потенциала способны освоить весь земной шар за несколько часов. Крупным организмам с более низким потенциалом для этого потребовалось бы несколько десятилетий или даже столетий.
Обычно лимитирующее давление среды играет в динамике популяции роль отрицательной обратной связи, поэтому в уравнении динамики популяции появляется дополнительный сомножитель или слагаемое:
dN/dt = rN(1 - N/K),
где К - верхняя асимптота, соответствующая верхнему пределу численности популяции после того, как в системе наступит равновесие (ее величина определяется средой, точнее, надсистемой).
В данном случае скорость роста равна максимальной скорости роста (rN), определяемой биотическим потенциалом, умноженной на степень реализации максимальной скорости (1 - N/K), которая тем ниже, чем больше плотность, или численность N, популяции.
Простейшей разностной функцией, удовлетворяющей таким условиям, является зависимость, предложенная Мальтусом: Xi+1 = RXi(1 - Xi). Это так называемое разностное логистическое уравнение.
Как и в предыдущем уравнении величина R является коэффициентом роста.
Новый член (1 - Xi) удерживает рост в определенных границах, так как когда Xi возрастает 1 - Xi уменьшается. Для простоты анализа эту функцию мы записали в абстрактном виде. Величины Xi лежат в интервале от 0 до 1. Xi=0 означает полное вымирание, а Xi=1 наибольшую предельно достижимую численность данного вида особей.
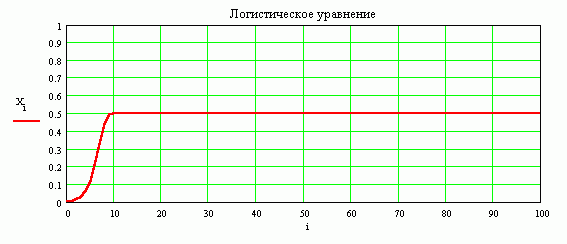
График решения при R = 2, начальное условие Xo = 0.004. Численность популяции первоначально мала. Вначале происходит медленный рост. Потом скорость увеличения численности резко возрастает и затем медленно приближается к стационарному значению.
![]()

График решения при R = 2.9, начальное условие Xo = 0.0009. Численность популяции первоначально мала. За счет большего параметра роста R, популяция быстро растет. Превышает стационарное значение, становиться меньше, затем снова значение численности возрастает, убывает и т. д. Однако, эти колебания имеют затухающий характер. С течением времени, достигнув стационарного значения численность популяции уже не меняется.

График решения при R = 3.4, начальное условие Xo = 0.0009. Здесь значение параметра роста R больше чем в предыдущем случае. Численность популяции в процессе развития во времени не достигает стационарного значения. Она все время регулярно меняется и принимает только два значения: наибольшее и наименьшее.
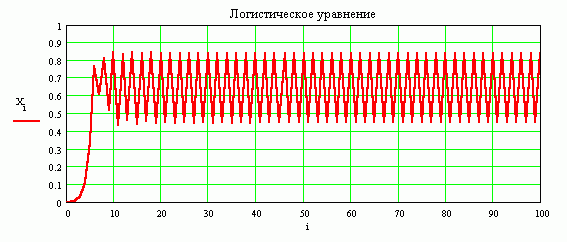
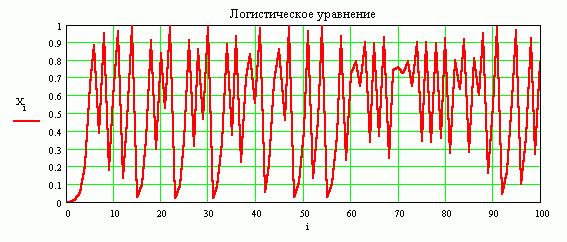
График решения при R = 3.99, начальное условие Xo = 0.0009. При том же значении начальной численности популяции Xo, величина параметра R роста очень велика и близка к максимальному значению 4 для модели. В такой ситуации, на графиках не видно стационарного значения, также не наблюдается повторяемость решений с течением времени развития популяции.
11.Модель конкуренции. Система уравнений Вольтерры–Лотки. Их графическая интерпретация. Поведение модельной системы и исход конкуренции в зависимости от соотношения параметров уравнений.
Межвидовая конкуренция
- это любое взаимодействие между двумя или более популяциями, которое отрицательно сказывается на их росте и выживании. Независимо от того, что лежит в основе межвидовой конкуренции - использование одних и тех же ресурсов или хищничество, - она может привести либо к взаимному приспособлению видов, либо к вытеснению одного вида другими.
Два видов, конкурирующих за один ресурс не могут устойчиво сосуществовать в гомогенной среде. При этом:
один вид вытесняется другим;
происходит смещение признаков (расхождение видов по экологическим особенностям).
Механизмы выхода из конкуренции:
пространственное расхождение (топическая специализация - в пространстве);
временн’ое расхождение (по времени, или во времени); Ночные, дневные, сумеречные
трофические предпочтения (трофическая специализация); Полифагия, олигофагия
поведенческие отличия (этологическая специализация); Спаривание, ухаживание
При наличии конкуренции скорость роста каждой популяции равна нелимитируемой скорости за вычетом величины, характеризующей влияние собственного перенаселения, которое возрастает с увеличением численности популяции, величины, характеризующей вредное влияние другого вида, которое с увеличением численности обоих видов N1 и N2 также возрастает:
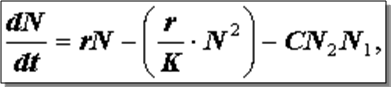
![]()
где N1 и N2 - численность взаимодействующих популяций.
В данном уравнении выражение dN/dt означает скорость роста; rN - нелимитированную скорость;
![]() -
влияние роста численности;
-
влияние роста численности;
CN2N1 - повреждающее влияние другого вида
Взаимодействие между двумя популяциями может привести к различным последствиям.
Если величина С для обоих видов незначительна, то подавляющее влияние со стороны другого вида будет иметь меньшее значение, чем внутривидовая конкуренция.
Если значение С велико, вид, оказывающий наибольшее влияние, может уничтожить своего конкурента или вынудит его покинуть данную территорию,
При благоприятном взаимодействии видов влияющих друг на друга популяций в уравнение вводится CN2N1 с положительным знаком. В этом случае популяции растут и процветают, достигая равновесных значений.
Законы конкуренции Лотки – Вольтерра
Для моделирования конкуренции типа хищник - добыча, паразит - хозяин или других видов взаимодействия в ограниченном пространстве можно использовать следующие уравнения, предложенные Лоткой (Lotka, 1925) и Вольтеррой (Volterra, 1926), полученные ими независимо друг от друга.
Рассмотрим уравнение логистического роста популяции:
![]() (1)
(1)
При этом, как известно, выражение в скобках отражает результат внутривидовой конкуренции.
В основе уравнения Лотки-Вольтерра лежит замена этого выражения таким, которое учитывает внутри и межвидовую конкуренцию.
Обозначим численность популяции первого вида N1, второго N2, предельную плотность насыщения и максимальную скорость роста как K1, K2, r1 и r2.
Предположим, что 10 особей вида 2 оказывают воздействие на вид 1, такое же как только 1 его особь. То есть, мы можем ввести коэффициент пересчета влияния особей одного вида в особи другого вида. К примеру, в данном случае это коэффициент будет равен 1/10.
Таким образом, совместное влияние (внутри- и межвидовой) конкуренции на вид 1 можно представить:
![]() (2)
(2)
При этом, константа 1/10 обозначается как α12.
С помощью коэффициента α12 оценивают конкурентное воздействие вида 2 на вид 1, в расчете на 1-у особь.
Умножая (N2) на (a12) мы выражаем его эквивалентным числом особей (N1).
При этом мы имеем следующее:
если
α12 < 1, то вид 2 оказывает меньшее ингибирующее воздействие на вид 1, чем вид 1 сам на себя, то есть внутривидовая конкуренция выше, чем межвидовая.
α12 > 1, то вид 2 оказывает большее влияние на вид 1, чем вид 1 влияет на себя, то есть межвидовая конкуренция выше, чем внутривидовая.
α12=α21, то конкуренты угнетают друг друга в равной степени.
Таким образом, если в выражение (1), вместо N, подставить выражение (2), то получим уравнение следующего вида:
![]()
и для 2-го вида соответственно
![]()
где
К1, K2 - предельная плотность при раздельном существовании;
N1, N2 - численность первого и второго видов;
r1 , r2 - максимальная скорость роста первого второго видов;
альфа - коэффициент, характеризующие тормозящее влияние вида (2) на вид (1) - (а12)
бетта - коэффициент, характеризующий тормозящее влияние вида (1) на вид (2) - (а21)
В рамках вольтерровской модели конкуренции результат конкуренции определяется следующими соотношениями:
a12 < K1 / K2 , a21 > K2 / K1 , выживает только вид i = 1 ;
a12 > K1 / K2 , a21 < K2 / K1 , выживает только вид i = 2 ;
a12 > K1 / K2 , a21 > K2 / K1 , выживает один или другой вид в зависимости от их начальных плотностей;
a12 < K1 / K2 , a21 < K2 / K1 , выживают оба вида.
Ki - максимально допустимая плотность популяции (емкость экологической ниши);
aij - коэффициенты конкуренции.
Теоретически два вида могли бы существовать на одной и той же территории, если бы характерные для них коэффициенты конкуренции были не очень велики по сравнению с величинами отношений плотностей насыщения.
12. Хищничество и его типы. Функциональная реакция потребителя на увеличение числа жертв. Математическая модель Лотки – Вольтерры.
Хищничество– кратковременный контакт, когда один организм вызывает гибель другого.:
Таксономическая классификация – плотоядные, растительноядные, всеядные организмы;
Функциональная классификация – истинные хищники, хищники с пастбищным типом питания, паразитоиды (сходные образ жизни), паразиты.
Отличия хищничества от паразитизма: хищник убивает добычу, паразит использует ее как среду обитания (более специализирован).
Существуют две классификации хищников. По одной из них (функциональной) выделяют 4 типа хищников: истинные хищники, хищники с пастбищным типом питания, паразиты и паразитоиды.
Истинные хищники убивают жертву почти сразу после нападения. В течение жизни хищники уничтожают довольно много разных особей. Часто они поедают жертву целиком, некоторые - только ее часть.
Хищники с пастбищным типом питания в течение жизни также убивают большое число жертв, но поедают их только частично. Воздействие их на жертву может быть различным, но чаще вредным- Нападение хищника на жертву редко приводит к ее скорой гибели. Точно предсказать летальный исход почти невозможно. К хищникам такого типа относятся крупные травоядные животные.
Паразиты, подобно хищникам с пастбищным типом питания, поедают только часть своей жертвы (хозяина). Их нападение, как правило, причиняет вред, но редко приводит к гибели жертвы. В отличие от хищников с пастбищным типом питания паразиты в течение жизни нападают на одну или ограниченное число жертв. Паразиты могут нападать не только на животных, но и на растения.
Паразитоиды - группа насекомых, выделенная на основе поведения взрослых самок при откладке яиц и личинок. Взрослые насеко-мые-паразитоиды ведут свободный образ жизни, но яйца откладывают либо в тело другого насекомого, либо на его поверхность. Вылупившиеся из яйца личинки паразитоидов развиваются внутри своего хозяина, который обычно сам не достиг взрослого состояния. Вначале личинка наносит небольшой вред хозяину, но по мере развития она его уничтожает.
Эволюция (изменение численности, выработка морфологических, физиологических и поведенческих адаптаций) жертвы и хищника происходит параллельно.
Хищничество, паразитизм и конкуренция способствуют регуляции численности и формированию видового состава. Хищничество и паразитизм обеспечивают круговорот веществ по пищевым цепям.
Согласно модели Лотки-Вальтера, колебания численности хищников идут параллельно с колебаниями численности жертв. Если число жертв увеличивается, то: 1) увеличивается число жертв (съеденных) за единицу времени; 2) увеличится численность популяции хищников:
Отличия хищничества от паразитизма: хищник убивает добычу, паразит использует ее как среду обитания (более специализирован).
Эволюция (изменение численности, выработка морфологических, физиологических и поведенческих адаптаций) жертвы и хищника происходит параллельно.
Паразитами являются 10% живых организмов. Паразитизм может быть постоянным (гельминт), временным (комар) или периодическим (личинка). Паразитизм также бывает обязательный и факультативный.
Функциональная реакция заключается в том, что каждый хищник начинает «лучше питаться», т.е. добывать большое число жертв. Однако съесть больше определенного предела он не может, поэтому если жертвы в благоприятных условиях размножаются слишком быстро, хищники уже не регулируют их численность. Но через некоторое время следует количественная реакция – увеличение числа самих хищников за счет успешного размножения при наличии хорошей кормовой базы. Это приводит к возрастанию потребления жертв уже в геометрической прогрессии. Поэтому после некоторого всплеска численности жертвы вновь попадают под регулирующее воздействие потребителей.
13.Паразитизм. Классификация форм паразитизма. Размещение паразитов. Конкуренция между паразитами. Мутуализм и его типы. Примеры мутуализма. Классификация мутуализма по уровню взаимодействия структур