

1. Кепирование 100% mРнк
2. Полиаденилирование ~95% mРНК
3.Сплайсинг ~95% mРНК. Сплайсингу подвергаются только полиаденилированные mРНК.
4.Редактирование Показано лишь для нескольких mРнк.
Все стадии процессинга mРНК происходят в РНП-частицах (рибонуклеопротеидных комплексах.
По мере синтеза про-mРНК, она тут же образует комплексы с ядерными белками - информоферами. И в ядерные, и в цитоплазматические комплексы mРНК с белками (информосомы) входят sРНК.
![]()
Таким образом, mРНК не бывает свободной от белков.
![]()
На всем пути следования до завершения трансляции mРНК защищена от нуклеаз. Кроме того, белки придают ей необходимую конформацию.
![]()
Определение: полисома - комплекс mРНК с несколькими или многими рибосомами.
![]()
В составе информосом mРНК может жить от нескольких минут до нескольких дней, не подвергаясь действию нуклеаз. (Так, mРНК живут неделями в ооцитах, предшественниках яйцеклеток).
![]()
Кепирование
Кепирование - надевание "шапочки".
|
|
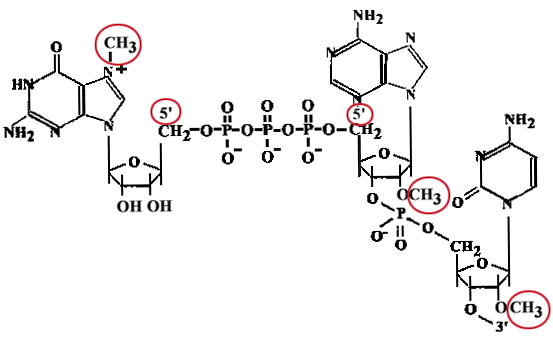
"Сар" представляет собой метилированный ГТФ, присоединенный в необычной позиции 5'-5' и две метилированные рибозы в первых двух нуклеотидах mРНК. По мере образования про-mРНК (еще до 30-ого нуклеотида), к 5'-концу, несущему пуринтрифосфат, присоединяется гуанин, после чего происходит метилирование.
ГТФ + гуанинтрансфераза
(Е)![]() Е~фГ + ф-ф
Е~фГ + ф-ф
5'ф-ф-ф-Пур-ф-Х-ф-Y-ф-Z-...+E~фГ![]() Г5'-ф-ф-ф-5'-Пур-ф-Х-ф-Y-ф-Z...
Г5'-ф-ф-ф-5'-Пур-ф-Х-ф-Y-ф-Z...![]()
![]() (метилирование)
Гm-ф-ф-ф-Пурm-ф-Хm-ф-Y-ф-Z-...
(метилирование)
Гm-ф-ф-ф-Пурm-ф-Хm-ф-Y-ф-Z-...
Назначение "Сар"
1. Защита 5'-конца mРнк от действия экзонуклеаз.
2. За счет узнавания "Сар"-связывающими белками происходит правильная установка mРнк на рибосоме.
![]()
Полиаденилирование
|
|
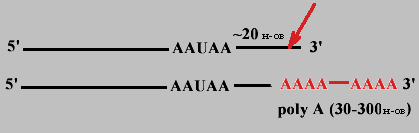
Когда синтез про-mРНК завершен, то на расстоянии примерно 20 нуклеотидов в направлении к 3' - концу от последовательности 5'-AAУAA-3' происходит разрезание специфической эндонуклеазой и к новому 3'-концу присоединяется от 30 до 300 остатков АМФ (безматричный синтез).
Каждый вид mРНК имеет "поли-А хвост" определенной длины. Он защищает 3'-конец от гидролиза, т.к. покрыт полиА-связывающими белками.
В значительной степени время жизни mРНК коррелирует с длиной полиА-хвоста.
![]()
mРНК ряда генов не полиаденилируется (например гистоновых генов).
Полиаденилированные про-mРНК подвергаются сплайсингу.
![]()
Сплайсинг
В 1978г. Филипп Шарп (Массачусетский технологический институт) открыл явление сплайсинга РНК (от англ. to splace - сшивать без узлов).
![]()
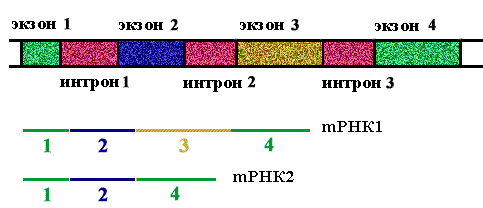
Определение: экзоны - кодирующие участки генов.
Определение: интроны - некодирующие участки генов.
![]()
На долю интронов приходится в 5-7 раз больше нуклеотидных пар, чем на долю экзонов. Количество экзонов в гене больше, чем интронов.
![]()
|
|
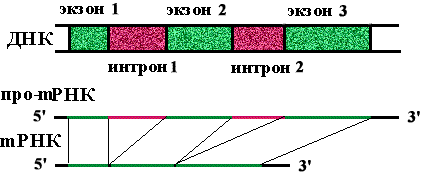
Определение: сплайсинг - вырезание копий интронов из про-mРНК и сшивание копий экзонов с образованием mРНК.
![]()
Копии интронов гидролизуются до нуклеотидов.
Сплайсинг показан для большинства mРНК и некоторых tРНК. У простейших найден автосплайсинг rРНК. Сплайсинг показан даже для археобактерий.
Не существует единого механизма сплайсинга. Описано по крайней мере 5 разных механизмов.
В ряде случаев сплайсинг осуществляют ферменты-матюразы.
В некоторых случаях в процессе сплайсинга участвуют sРНК.
В случае автосплайсинга процесс происходит благодаря третичной структуре про-РНК.
Для mРНК высших организмов существуют обязательные правила сплайсинга:
![]()
Правило 1. 5' и 3' концы интрона очень консервативны: 5'(ГT-интрон-AГ)3' .
|
|
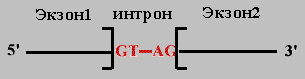
Правило 2. При сшивании копий экзонов соблюдается порядок их расположения в гене, но могут быть выброшены некоторые из них.
|
|
![]()
Представление об интроне, как пустой, ничего не кодирующей последовательности, неверно. Некоторые интроны кодируют ферменты-матюразы, вырезающие копии этих интронов. На вопрос, зачем эукариотическим геномам экзон - интронная структура, можно ответить только в единичных случаях. Почему эукариотические гены "разорваны", будет рассмотрено в конце курса.
![]()