

Как образуются рибосомы у эукариот
Гены rРНК присутствуют в количестве от 10 до 105 копий у разных видов (105 у амфибий). У человека - 300 генов, в которых закодированы rРНК.
Все рибосомные гены, кроме генов 5S рибосомной РНК, сближены (т.е располагаются один за другим) и образуют несколько кластеров. Сначала синтезируется про-rРНК, после созревания которой образуются 28S, 18S и 5,8S rРНК.
|
|

Интерфазные хромосомы в световой микроскоп не видны. Каждый ген прорибосомной РНК транскрибируется одновременно несколькими РНК-полимеразами и тут же начинается процессинг.
На электронномикроскопических фотографиях видна картина "рождественской елочки". Синтезируемые в ядре mРНК поступают на готовые рибосомы в цитоплазму, где синтезируются рибосомные белки, которые идут в ядро и путаются в "ветвях елки".
Образуются рибосомные субъединицы. Одновременно в эукариотическом ядре находятся сотни тысяч субъединиц рибосом.
![]()
Определение: ядрышко - место образования субъединиц рибосом, наблюдаемое в световой микроскоп.
![]()
В ядре может быть несколько ядрышек.
Определение: кластер генов rРНК называют ядрышковым организатором.
Особенности транскрипции эукариот
Единицей транскрипции у эукариот является отдельный ген, а не оперон, как у прокариот.
Оператор, как таковой, отсутствует.
Промотор есть, но он организован иначе.
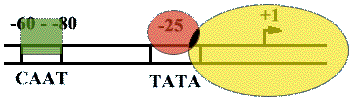
На расстоянии -25 п.н. от +1 нукл. находится ТАТА-бокс. Его позиция определяет точку инициации транскрипции. А на расстоянии -60-80 п.н. находится ЦААТ-бокс, который не является абсолютно необходимым, но присутствует перед большинством генов. Расстояние между ЦААТ и ТАТА большое и РНК-полимераза не способна накрыть всю эту область. ЦААТ опознается своим белком, а ТАТА - своим.
|
|
Помимо этих есть еще несколько белков, называемых базальными факторами транскрипции.
![]()
Определение: базальные факторы транскрипции - белки, необходимые для инициации транскрипции.
![]()
Базальные факторы транскрипции необходимы для инициации транскрипции всеми тремя ядерными РНК-полимеразами.
Для любого гена, кодирующего белок, есть энхансеры (усилители).
![]()
Определение: энхансеры - последовательности ДНК, усиливающие транскрипцию при взаимодействии со специфическими белками.
![]()
|
Энхансеры - это не непрерывные последовательности нуклеотидов. Существуют так называемые модули - это отдельные части энхансеров. Одинаковые модули могут встречаться в разных энхансерах. Для каждого энхансера набор модулей уникален. Модули - это короткие последовательности, не более 2-х витков спирали (20 п.н.), которые могут находиться перед, за и даже внутри гена. |
|
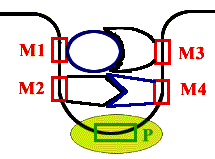
Таким образом, М1+М2+М3+М4 - один энхансер, но он состоит из 4-х модулей. Все 4 модуля узнаются своими белками, а они, сидя на ДНК, взаимодействуют друг с другом. Если в клетке присутствуют все соответствующие белки, то участку ДНК придается определенная конформация и начинается синтез mРНК.
![]()
Все соматические клетки многоклеточного эукариотического организма имеют абсолютно одинаковый набор генов. Почему же клетки дифференцированы и специализированы?
Дело в том, что все гены работают на фоновом уровне и не имеют фенотипического проявления. Экспрессируются лишь те гены, у которых все энхансерные модули узнаны своими белками и эти белки взаимодействуют друг с другом. Кроме энхансеров есть сайленсеры (ослабители).
![]()
Определение: сайленсеры - последовательности ДНК, ослабляющие транскрипцию при взаимодействии с белками.
![]()
При соответствующем наборе белков экспрессия отдельных генов в клетке может быть подавлена.
Процессинг (созревание) rРНК и tРНК у эукариот принципиально не отличается от такового у прокариот.
Процессинг mРНК отличается сильно и состоит из нескольких этапов.