

2.1.3.Механизм проведения возбуждения по нервному волокну
Биопотенциалы могут быть локальными (местными), распространяющимися с декрементом (затуханием) на расстояние, не превышающее 1—2 мм, и импульсными (ПД), распространяющимися без декремента по всей длине волокна — на несколько десятков сантиметров, например от мотонейронов спинною мозга по всей длине нервного волокна до мышечных волокон конечностей с учетом длины самой конечности.
Распространение локальных потенциалов. Локальные потенциалы (препотенциал, рецепторный потенциал, возбуждающий постсинаптический потенциал — ВПСП) изменяют мембранный потенциал покоя, как правило, в сторону деполяризации в результате входа в клетку Na+ согласно электрохимическому градиенту. В результате этого между участком волокна, в котором возник локальный потенциал, и соседними участками мембраны формируется электрохимический градиент, вызывающий передвижение ионов. В частности, вошедшие в клетку ионы Na+ начинают перемещаться в соседние участки, а ионы Na+ на наружной поверхности клетки движутся в противоположном направлении. В итоге поляризация мембраны соседнего участка уменьшится. Фактически это означает, что локальный потенциал из первичного очага распространился на соседний участок мембраны. Он затухает на расстоянии 1—2 мм от очага первичной деполяризации, что связано с отсутствием ионных управляемых каналов на данном участке мембраны или неактивацией управляемых ионных каналов, продольным сопротивлением цитоплазмы волокна и шунтированием тока во внеклеточную среду через каналы утечки мембраны.
Если
возникшая деполяризация мембраны не
сопровождается изменением проницаемости
потенциалзависимых натриевых, кальциевых
и калиевых каналов, такую деполяризацию
называют электротонической.
Электротоническое
распространение возбуждения —
физический механизм, оно характерно
для
тех фрагментов мембран возбудимых
клеток,
где нет потенциалзависимых ионных
каналов.
Такими участками являются, например,
большая часть мембраны дендритов нервных
клеток, межперехватные промежутки
в миелиновых нервных волокнах. Если
местный
потенциал (рецепторный или ВПСП),
распространяясь электротонически,
достигает
участков мембраны, способных генерировать
ПД (аксонный холмик, перехваты
Ранвье, часть мембраны дендритов и,
возможно,
сомы), но его амплитуда при этом не
достигнет критического уровня
деполяризации,
то такой потенциал называют препотенциалом.
В его возникновении и распространении
частично участвуют потенциалзависимые
ионные каналы, однако при этом нет
регенеративной (самоусиливающейся)
деполяризации,
характерной для ПД. Поэтому распространение
такого потенциала происходит с
затуханием амплитуды. Если локальный
потенциал
достигает участков мембраны, способных
генерировать ПД, и его амплитуда выходит
 на
критический уровень деполяризации,
формируется ПД, который распространяется
по всей длине нервного волокна без
затухания.
на
критический уровень деполяризации,
формируется ПД, который распространяется
по всей длине нервного волокна без
затухания.
Эффективность электротонического распространения биопотенциалов зависит от физических свойств нервного волокна — сопротивления и емкости его мембраны, сопротивления цитоплазмы. Электротоническое проведение в нервном волокне улучшается при увеличении его диаметра, что связано с уменьшением сопротивления цитоплазмы, а также при миелинизации волокна, увеличивающей сопротивление мембраны (до 105 Ом/см2) и уменьшающей ее емкость (до 0,005 мкФ/см2). Эффективность электротонического проведения характеризует постоянная длины мембраны (λm). Это расстояние, на которое может электротонически распространиться биопотенциал, пока его амплитуда не уменьшится до 37 % от исходной величины. Постоянная длины для локальных потенциалов реально не превышает 1 мм, и их амплитуда затухает на расстоянии 1—2 мм от места возникновения.
Для передачи возбуждения на большие расстояния необходимо формирование ПД. В его распространении, кроме электротонического механизма, существенная роль принадлежит механизму регенеративной деполяризации, позволяющей сохранить амплитуду ПД на всем пути его следования.
Проведение потенциала действия. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов, ответственных за формирование ПД. В распространении ПД можно выделить два этапа: этап электротонического проведения, обусловленный физическими свойствами нервного волокна, и этап генерации ПД в новом участке на пути его движения, обусловленный реакцией ионных каналов. В зависимости от расположения и концентрации ионных каналов в мембране волокна возможно два типа проведения ПД: непрерывный и сальтаторный (скачкообразный).
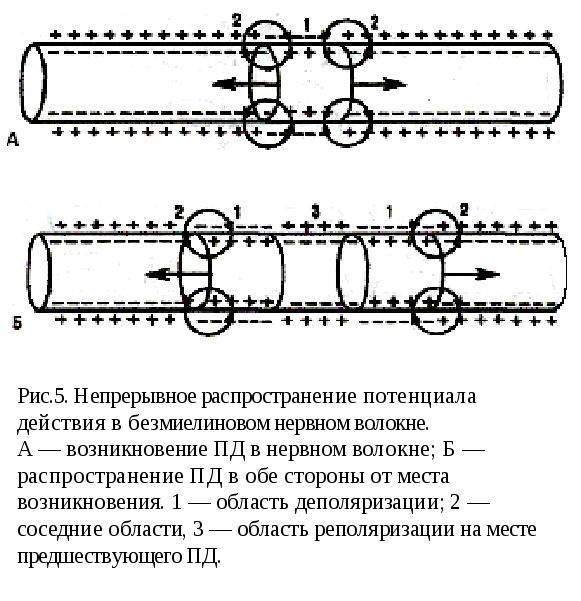
Непрерывное распространение ПД осуществляется в безмиелиновых волокнах типа С, имеющих равномерное распределение потенциалзависимых ионных каналов, участвующих в генерации ПД. Проведение нервного импульса начинается с этапа электротонического распространения возникшего ПД. Амплитуда ПД нервного волокна (мембранный потенциал + инверсия) составляет около 90 мВ, постоянная длины мембраны (λm) в безмиелиновых волокнах равна 0,1—1,0 мм.
Поэтому ПД, распространяясь на этом расстоянии как электротонический потенциал и сохранив как минимум 37 % своей амплитуды, способен деполяризовать мембрану до критического уровня и генерировать на всем протяжении новые ПД (рис.5). При этом на этапе электротонического распространения нервного импульса ионы движутся вдоль волокна между деполяризованным и поляризованным участками, обеспечивая проведение возбуждения в соседние участки волокна. Реально при неповрежденном нервном волокне этап чисто электротонического распространения ПД (вдоль мембраны) предельно мал, так как потенциалзависимые каналы имеются в непосредственной близости друг от друга и, естественно, — от возникшего потенциала действия и наблюдается только до достижения деполяризации, равной 50 % Екр. Далее включается перемещение ионов в клетку (нервное волокно) и из клетки за счет активации ионных т каналов.
При формировании нового ПД в соседнем участке в фазе деполяризации возникает мощный ток ионов натрия в клетку вследствие активации натриевых каналов, приводящий к регенеративной (самоусиливающейся) деполяризации. Этот ток обеспечивает формирование нового ПД той же амплитуды, представляющий собой, как обычно, сумму двух величии н — мембранного потенциала покоя и инверсии. В связи с этим проведение ПД осуществляется без декремента (без снижения амплитуды). Таким образом, непрерывное распространение нервного импульса идет через генерацию новых ПД по эстафете, когда каждый участок мембраны выступает сначала как раздражаемый (при поступлении к нему электротонического потенциала), а затем как раздражающий (после формирования в нем нового ПД).
С альтаторный
( прерывистый, скачкообразный )
тип
проведения
нервного
импульса осуществляется в миелиновых
волокнах (типы А и В), для которых
характерна
концентрация потенциалзависимых ионных
каналов только в небольших участках
мембраны (в перехватах Ранвье), где их
плотность достигает 12 000 на 1 мкм2, что
примерно в 100 раз выше, чем в мембранах
безмиелиновых
волокон. В области миелиновых муфт
(межузловых сегментов), обладающих
хорошими изолирующими свойствами,
потенциалзависимых
каналов почти нет, и мембрана
осевого цилиндра там практически
невозбуд
има. В этих условиях ПД, возникший
в одном перехвате Ранвье, электротонически
(вдоль волокна, без участия ионных
каналов)
распространяется до соседнего перехвата,
деполяризуя там мембрану до критического
уровня, что приводит к возникновению
нового ПД, т.е. возбуждение проводится
скачкообразно (рис. 6). Постоянная длина
мембраны миелинового волокна достигает
5 мм. Это значит, что ПД, распространяясь
электротонически на этом расстоянии,
сохраняет 37 % своей амплитуды (около
30 мВ) и может деполяризовать мембрану
до критического уровня (пороговый
потенциал в перехватах Ранвье равен
около 15
мВ). Поэтому в случае повреждения
ближайших
на пути следования перехватов Ранвье
потенциал действия может электротонически
возбудить 2—4-й и даже 5-й перехваты.
Сальтаторное
проведение ПД по миелиновым
волокнам является эволюционно более
поздним
механизмом, возникшим впервые у
позвоночных.
Оно имеет два важных преимущества
по сравнению с непрерывным проведением
возбуждения. Во-первых,
оно
более экономично в энергетическом
плане, т.к. возбуждаются
только перехваты Ранвье, площадь которых
менее 1 % мембраны, и, следовательно,
надо меньше энергии для восстановления
трансмембранных градиентов Na+
и К+,
уменьшающихся
в процессе формирования ПД.
Во-вторых,
возбуждение
проводится с большей
скоростью (см. табл.3), чем в безмиеликовых
волокнах, так как возникший ПД на
протяжении миелиновых муфт распространяется
электротонически, что в 107
раз быстрее,
чем скорость непрерывного проведения
ПД
в безмиелиновом волокне.
альтаторный
( прерывистый, скачкообразный )
тип
проведения
нервного
импульса осуществляется в миелиновых
волокнах (типы А и В), для которых
характерна
концентрация потенциалзависимых ионных
каналов только в небольших участках
мембраны (в перехватах Ранвье), где их
плотность достигает 12 000 на 1 мкм2, что
примерно в 100 раз выше, чем в мембранах
безмиелиновых
волокон. В области миелиновых муфт
(межузловых сегментов), обладающих
хорошими изолирующими свойствами,
потенциалзависимых
каналов почти нет, и мембрана
осевого цилиндра там практически
невозбуд
има. В этих условиях ПД, возникший
в одном перехвате Ранвье, электротонически
(вдоль волокна, без участия ионных
каналов)
распространяется до соседнего перехвата,
деполяризуя там мембрану до критического
уровня, что приводит к возникновению
нового ПД, т.е. возбуждение проводится
скачкообразно (рис. 6). Постоянная длина
мембраны миелинового волокна достигает
5 мм. Это значит, что ПД, распространяясь
электротонически на этом расстоянии,
сохраняет 37 % своей амплитуды (около
30 мВ) и может деполяризовать мембрану
до критического уровня (пороговый
потенциал в перехватах Ранвье равен
около 15
мВ). Поэтому в случае повреждения
ближайших
на пути следования перехватов Ранвье
потенциал действия может электротонически
возбудить 2—4-й и даже 5-й перехваты.
Сальтаторное
проведение ПД по миелиновым
волокнам является эволюционно более
поздним
механизмом, возникшим впервые у
позвоночных.
Оно имеет два важных преимущества
по сравнению с непрерывным проведением
возбуждения. Во-первых,
оно
более экономично в энергетическом
плане, т.к. возбуждаются
только перехваты Ранвье, площадь которых
менее 1 % мембраны, и, следовательно,
надо меньше энергии для восстановления
трансмембранных градиентов Na+
и К+,
уменьшающихся
в процессе формирования ПД.
Во-вторых,
возбуждение
проводится с большей
скоростью (см. табл.3), чем в безмиеликовых
волокнах, так как возникший ПД на
протяжении миелиновых муфт распространяется
электротонически, что в 107
раз быстрее,
чем скорость непрерывного проведения
ПД
в безмиелиновом волокне.