

6.Отличия репликации у эукариот и прокариот
Отличия репликации у эукариот связаны с нуклеосомным строением хромосом. ДНК наматывается на частички нуклеосом и межнуклеосомные фрагменты ДНК включают примерно 200 нуклеотидов. Поэтому фрагменты Оказаки у эукариот короче, чем у прокариот и составляют - 100-200 нуклеотидов. В связи с нуклеосомным строением меньше и скорость синтеза ДНК (50-100 нуклеотидов в сек). Нуклеосомы перед репликацией распадаются и формируются заново на новых цепях на расстоянии 200-400 нуклеотидов от репликативной вилки. Репликация идет в двух взаимопротивоположных направлениях с множеством репликонов. У эукариот не 1 репликон, как у прокариот, а множество: у дрожжей – 500, у млекопитающих – 20-30 тыс.
Типы репликации
Репликация у всех исследованных на данный момент организмов репликация происходит подобным образом, однако некоторые различия имеются в связи с различной формой хромосом. Различают 3 основных типа репликации: Ϋ-тип (игрек), θ-тип (тэта), σ-тип (сигма). Ϋ-тип репликации характерен для линейных хромосом эукариот и некоторых вирусов, так как у них репликативная вилка напоминает букву “игрек”. Репликация кольцевых хромосом бактерий, плазмид, фага λ сопровождается локальным расхождением цепей кольцевой ДНК в oriCи напоминает греческую букву «тэта», поэтому и получила названиеθ-типа. σ-тип репликации характерен для вирусов с одноцепочечной ДНК или РНК и напоминает катящееся кольцо или разворачивающийся рулон. При этом матричная нуклеиновая кислота напоминает вращающееся кольцо, вокруг которого синтезируется дочерняя цепь. Причем сформированная дочерняя цепочка может содержать несколько тандемно расположенных копий генома вируса, которые впоследствии “разрезаются” специальным ферментом.
8. Проблема репликации линейных концов днк. Теломераза.
После окончания репликации у эукариот с (5`)-концов происходит гидролиз РНК-овых транскриптов, в результате чего дочерние цепи оказываются короче материнских 3`-концов цепей. В 1998г. В журнале «Наука» («Science») Джери Шей опубликовалстатью оботкрытии фермента –теломеразы, способном производитьудлинение недостающего фрагмента, т.е.синтезировать теломеры –концы хромосом,состоящие изповторяющихся блоков ТТАГГГ. Оказалось,что этот ферментработает постоянно в ракових клетках иобеспечивает их потенциальное бессмертие. Также он был обнаружен в половыхи стволовых клетках, обеспечивающих обновление клеток крови. В соматических клетках скаждой репликациейтеломеры уменьшаютсяприблизительно на 50нуклеотидов,т.к. неработает теломераза ииз-за этого примерно через 50 делений клетка гибнет.
Теломераза представляет собой нуклеопротеид и состоит из белковой глобулы и фрагмента РНК (у человека длиной 450 нуклеотидов). Теломераза присоединяется к концу ДНК и достраивает длинный 3`-конец, используя РНК, как матрицу, а затем на удлиненном конце осуществляется дополнительная репарирующая репликация с застраиванием уже 5`-конца.
Проблема изучения теломеразы очень важна для продления жизни людей и для решения проблемы рака. Продолжительность жизни животных, в клетки которых был введен ген теломеразы, увеличивалась на 40 %. Однако, без знания регуляторных факторов теломеразы ее использование может привести к злокачественному перерождению клеток. И наоборот, обнаружение ингибиторов теломеразы может дать в руки онкологов действенное оружие для борьбы с раком.
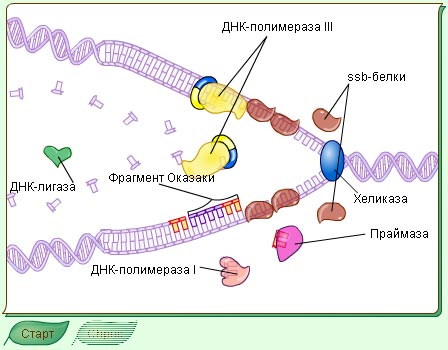
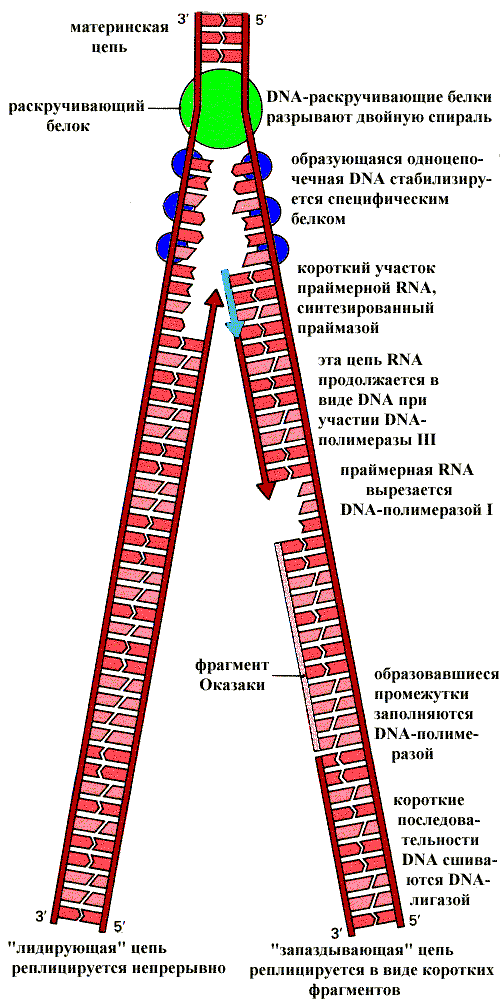
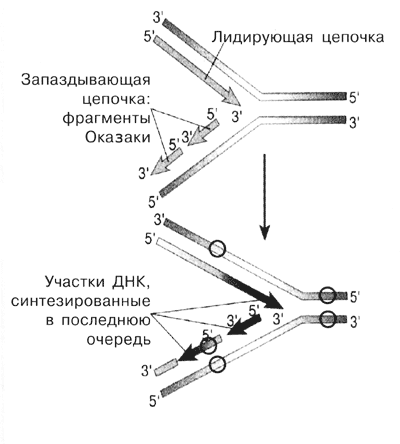
Рис.1-3. Репликация у прокариот на примере E.coli и участие в ней ферментов.Образование полуконсервативных "дочерних" молекул ДНК в результате репликации. Синтез "дочерних" цепей идёт в направлении от 5' к 3' комплементарно и антипараллельно цепям "материнской" молекулы
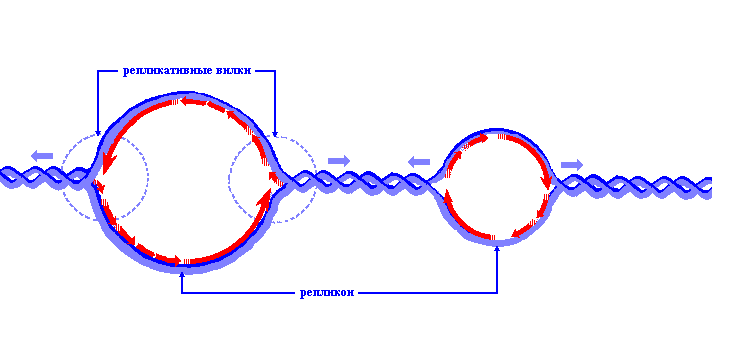
Рис.4. Репликация ДНК эукариотической хромосомы. Показан один из многих репликонов. Репликативные вилки движутся в противоположных направлениях от точки начала репликации.