

Метод дробовика (дробового ружья)
При невозможности синтезировать ген химическим путем или с помощью обратной транскриптазы используют технику Shot-gun– метод дробового ружья. Суть его заключается в том, что сначала ДНК дробят на фрагменты с помощью рестриктаз,то есть создают библиотку генов или фрагментов ДНК. Полученные ДНК-овые фрагменты гибридизируют «вслепую» с вектором, разрезаным той же рестриктазой. Гибридные векторы, содержащие встроенный ген методом трансформации переносят в бактерии (обычно в кишечную палочку) и размножают. Затем идентифицируют и отбирают необходимые колонии одним из перечисленных ниже методов (см.IVэтап). Способ отбора на селективных средах осуществляют с помощью генетических маркеров – способности расти на средах с антибиотиком, если ген несет фактор устойчивости, неспособности расти на средах с антибиотиком, если ген устойчивости к этому антибиотику инактивирован путем встройки в него гибридного фрагмента (при этом используется метод реплик), способности расти на средах без ауксина, если в геном ауксотрофа по этому ауксину введен нужный ген. Если же соответствующий генетический маркер не найден, используют метод гибридизации.
Іі этап. Конструирование рекомбинантных молекул с помощью векторов. Векторы и принципы их конструирования
Для введения чужеродной ДНК в клетки реципиентов используют так называемые векторы. Векторы– это молекулы ДНК способные переносить размножать и хранить генетическую информацию. Это своеобразные транспортные средства для переноса чужеродной информации (ДНК) в реципиентные клетки. Наиболее подходящие для выполнения этих функций являются небольшие по размерам молекулы ДНК плазмид, транспозонов, вирусов, фагов, митохондрий и хлоропластов. Одной из важных задач на ряду с созданием рекомбинантной ДНК, является их молекулярное клонирование.
Векторы должны обладать рядом свойств:
способностью автономно размножаться в клетке реципиента;
иметь селективный маркер (свойство, по которому проводится отбор);
содержать несущественные для размножения области ДНК, куда «безболезненно» может быть встроена чужеродная ДНК;
иметь минимальное количество сайтов узнавания для рестриктаз;
В природном виде плазмиды и другие векторы практически не используются. Их приходится перестраивать в зависимости от конктретных нужд. Векторы могут использоватся для размножения рекомбинантной ДНК, для переноса и интеграции в клетку хозяина и последующей экспрессии донорного гена. Большое количество векторов сконструировано на основе плазмид.
Наибольшее распространиение получил вектор рВR322, (4,3 т.п.н.) сконструирован на основе плазмидColE1,рSC101иR1drd. Вектор содержит два гена устойчивости к тетрациклину и ампицилину и сайты узнавания для рестриктазHindIII, SalI, BarnH1, EcoR1,Pst1 и др. Сайты узнавания для некоторых рестриктаз находятся в генах устойчивости к тетрациклину и ампицилину. Встраивание в них чужеродных генов приводит к появлению рекомбинантов, чувствительных к этим антибиотикам, то есть эти гены выступают в качестве селективных генетичекких маркеров.
Дж. Шелл показал что Ті – плазмидаизAgrobakteriumtumefaciensможет перходить в клетки корней высшего растения (в часности табака) и встраиваться в геном. Это создало предпосылки для передачи генов азотфиксации высшим растениям.
В качестве векторов часто используют также фаг М13, фаг λ и вирус SV-40. Для клонирования больших фрагментов чужеродных ДНК часто используюткосмиды – гибриды, созданные на основе плазмиды рВR322 иcos-сайтов фага λ.Cos-сайты - это одноцепоченые «липкие» концы, которые в естественных условиях соединяют несколько молекул фаговой ДНК в единый конкатемер. При созревании фага ферменты, катализирующие упаковку фаговой ДНК, узнают в составе конкатемера дваcos-сайта, выщепляют расположенный между ними фаговый геном и упаковывают его в головку фага. Точно также, введяcos-сайты в плазмиду рВR322, а именно, в ген устойчивости к ампициллину, а затем, разрезавcos-специфическим ферментомcos-сайт и чужеродную эукариотическую ДНК, можно получить короткие фрагменты этой эукариотической ДНК, упакованные в фаговые головки. Ими заражают кишечную палочку и отбирают рекомбинантные клетки только по устойчивости к тетрациклину, так как ген устойчивости к ампициллину нарушен. Космиды широко используют для создания банков генов.
Фазмидытакже являются гибридами фаговой и плазмидной ДНК. После встраивания в них чужеродных генов они в определенных условиях могут развиваться или как фаги, или как плазмиды.
Плазмиды и некоторые вирусные векторы оказались перспективными для внедрения генов в клетки больных страдающих наследственными заболеваниями и для лечения трудно излечимых болезней.
Малые векторы имеют то преимущество, что они многокопийны и с их помощью можно получить повышеный выход биотехнологического продукта.
Вектор и встраиваемая ДНК, разрезанная рестриктазами с образованием липких или тупых концов и после объединения фрагментов фосфодиэфирную связь между ними восстанавливают при помощи фермента ДНК-лигазы. Для сшивки тупых концов используют также РНК-лигазу фага Т4.
Для сшивки фрагментов с различающимися концами (тупыми и липкими, липкими с разной длиной) используют методы линкеров, коннекторов и адапторов.
Метод коннекторов состоит в том, что сшиваемые фрагменты ДНК удлиняются путем достраивания одноцепочечных концов ДНК с одной стороны тимидиловыми нуклеотидами (поли-Т), а с другой адениловыми нуклеотидами (поли-А). Таким образом у сшиваемых фрагментов ДНК появляются взаимно комплементарные цепи –коннекторы(соединители). Этот прием важен, если вектор и встраиваемую ДНК нельзя нарезать одной рестриктазой. В подобных случаях используют линкеры и адапторы – короткие синтетические или естественные фрагменты ДНК с одной стороны имеющие липкий конец, полученый при помощи одной рестриктазы, а с другой стороны – другой рестриктазы.
Метод линкеров. Линкеры– это химически синтезированные олигонуклеотиды, представляющие собой сайты рестрикции для эндонуклеаз (рестриктаз). Их достраивают с концов фрагментов ДНК, предназначенных для объединения. Затем обрабатывают всю ДНК рестриктазой, соответствующей линкеру, и получают фрагменты с комплементарными липкими концами. С помощью лигазы их сшивают. В результате получают рекомбинантные молекулы ДНК. Иногда в середину линкера встраивают какой-либо регуляторный участок, например, промотор гена или участок, обеспечивающий связывание с рибосомой (Шайна-дальгарно). Таким образом, линкеры не только способствуют объединению генов, но и обеспечивают их экспрессию в клетке реципиента.
Метод адапторов. Адапторы –это искусственно созданные фрагменты ДНК, которые можно использовать в виде своеобразных переходников. Один конец адаптора имеет конец комплементарный одному фрагменту сшиваемой ДНК, а другой конец адаптора комплементарен второму фрагменту. В адапторы также встраивают регуляторые элементы генов.
При необходимости липкие концы можно превратить в тупые, отщепляя выступающий липкий конец с помощью эндонуклеазы S1, которая разрушает только одноцепочечную ДНК. Иногда, наоборот, достраивают липкие концы, или застраивают бреши в одной из цепей ДНК с помощью ДНК-полимеразыI.
Для того, чтобы вскрытый вектор не смыкался без включения чужеродной ДНК, его обрабатывают фосфатазой, удаляющей фосфатной группы с 5' концов. В тоже время чужеродный фрагмент несет на 5' концах фосфатные группы, позволяющие при помощи лигазы замкнуть кольцо вектора.
рис.1
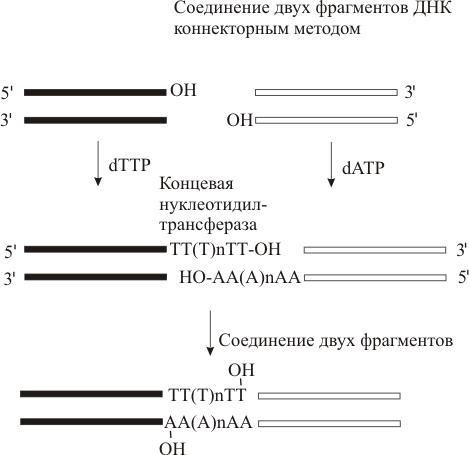
Рис. Коннекторый метод соединения концов ДНК