

BIO_-_vsyo
.pdf1. О сущности живого. Нуклеопротеидные комплексы.Эволюция представлений о химической сущности жизни.
Ф.Энгельс: «Жизнь – способ существования белковых тел» Жизнь – активная форма существования материи; период существования отдельно взятого организма от момента его возникновения до старости.
Нач XX в. академик Кольцов – гипотеза «Особых кольцевых молекул белков» ДНК как хим соед-е идентифицировано ещё в XIX в. Мишер.
Опыт Гриффитса 1926 – феномен трансформации (в феномене трансформации два участника: бакт и чужеродн ДНК, к-ая измен св-ва бактерии. ТФ – трансформирующий фактор - из убитого S-штамма вызвал превр-е R-штамма в S-штамм)
Гриффитс не смог определить химическую природу ТФ.
1944-лаб-я Эвери – экспериментальные доказательства - ТФ идентичен ДНК.
R + мышь – жив; S + мышь – мёртв; S(t) + мышь – жив; S(t) + R - мёртв
В живых системах 3 потока: ЭНЕРГИИ, ВЕЩЕСТВА и ИНФОРМАЦИИ, кот. подчиняются законам термодинамики.
1 ЗАКОН: В плане энергии нельзя выиграть (переходит из 1 вещ в другое)
2 ЗАКОН: В плане энергии нельзя остаться «при своих» (при переходе энергии ее часть теряется, выделяется в виде тепла)
Нуклеи к-ты (ДНК, РНК) и белки являются субстратом жизни. Ни нуклеин к-ты, ни белки в отдельности не являются субстратами жизни. Поэтому считают, что субстратами жизни являются нуклеопротеиды. Нет живых систем, не содержащих их (от вирусов до человека). Однако они являются субстратом жизни лишь когда находятся и функционируют в клетке, Вне клеток – это обычные химич соед-я. Следоват-но, жизнь – это взаимод-е нуклеин к-т и белков, а живое – то, что содержитсамовоспроизводящуюся
молекулярную систему в виде механизма активного воспроизв-ва синтеза нуклеиновых кислот и белков. Жизнь существует в виде нуклеопротеидных комплексов.
2. Клетка – миниатюрная биосистема. 5 признаков живых систем.
(см 1 вопр)
Клетка – это самостоятельная биосистема, уровень организации живой материи, кот присущи проявления основных свойств живого:
5 признаков живых систем:
1.Открытость (живые системы обмениваются с окр средой энергией, веществами, информацией)
2.Самообновление (системы эволюционируют во времени)
3.Саморегуляция (гомеостаз; системы не требуют регуляции из вне)
4.Самовоспроизведение
5.Высокоупорядоченность
Клетка представляет собой единицу строения, развития и размножения организмов - самоуправляемая система. Управляющая генетическая система клетки предствалена сложными макромолекулами - нуклеиновыми кислотами (ДНК и РНК). Клетка может существовать только как целостная система, неделимая на части. Целостность клетки обеспечивают биологические мембраны. Клетка - элемент системы более высокого ранга - организма. Части и органоиды клетки, состоящие из сложных молекул, представляют собой целостные системы более низкого ранга. Клетка рассматривается в качестве общего структурного элемента живых организмов.
Клеточня теория – одно из общепризнанных биологических обобщений, утверждающих единство принципа строения живых организмов.
Современная клеточная теория включает следующие основные положения:
1.Клетка – единица строения (все живые существа состоят из клеток).
2.Клетка – единица жизнедеятельности (все клетки сходны по строению, химическому составу и жизненным функциям).
3.Клетка – мельчайшая единица живого (каждая клетка реализует все св-ва живого)
4.Клетка – единица размножения (кажд клетка возникает из клетки) – Р.Вирхов
3.Клетка – элементарная еденица живого. Отличительные признаки
про- и эукариотических клеток.
Клетка – элементарная единица живого, основная единица строения, функционироваия, размножения и развития всех живых организмов. Клетка представляет собой биосистему, которой присущи все признаки живых систем.
Параметры сравнения |
Прокариоты (ядра нет) |
Эукариоты (есть ядро) |
Организмы |
Архебактерии, эубактерии (цианобактерии, зелёные |
Грибы, растения, животные |
|
синтезирующие бактерии;серные, метанообразующие) |
|
Размеры клетки |
1-10 мкм |
10-100 мкм |
Генетический материал |
2-х цепочечная Кольцевая молекула ДНК, |
Линейная ДНК организована с участиембольшого |
|
находящаяся в нуклеоиде и плазмидах. Отсутствуют |
кол-ва белков в хромосомы и заключена в |
|
белки-гистоны. Устойчив к антибиотикам. |
ядро;митохондрии и пластиды имеют собственную |
|
|
кольцевую ДНК. Есть белки-гистоны. |
Поверхностный аппарат |
Мембрана и надмембранные структуры (содерж |
Плазматич.мембрана, надмембр.и |
|
Муреина в клет стенке, преоблад белков над |
субмембр.комплекс(белки, фосфолипиды, |
|
липидами. Мезосома-впячивание мембраны внутрь |
полуинтегральные белки, гликокаликс,фурмент ф.-у |
|
для увелич поверхности. |
животных; у растений-целлюлоза). |
Цитоплазма |
Не разделена на компартменты, не содержит |
Есть цитоскелет, организующий цитоплазму и |
|
мембранных органоидов и волокон цитоскелета |
обеспеч.еёдвижение;находится много мембранных |
|
|
органелл. |
Немембр.структуры: |
|
|
Цитоскелет |
- |
+(микротрубочки, микрофиламенты, |
Рибосомы |
70S |
промежут.филаменты) |
|
|
80S(крупнее, чем ) |

Двумембр.стр-ры |
|
|
Митохондрии |
-(задатки. Вместо них-лизосомы) |
+(Имеют собственные рибосомы и кольцевую ДНК) |
Пластиды |
-(АТФ и фотосинтез-в растит кл.) |
+ |
Одномембр.стр-ры |
- (никаких нет) |
+ (всё есть) |
ЭПС |
|
|
Ап-т Гольджи |
|
|
Лизосомы |
|
|
Пероксисомы |
|
|
Вакуоли |
|
(в растительной клетке) |
Включения |
Белки+малые молек, зап питат вещ-ми |
капли жира, крахмал/гликоген |
Способ деления |
Бинарное деление, перетяжка, конъюгация. Амитоз. |
Митоз, мейоз, амитоз |
Движение |
Жгутик(из одного белка фибриллина) из белка- |
Жгутики, реснички, псевдоподии(у простейших) из |
|
флагмина |
белка-тобулина |
Особенности метаболизма |
Способность фиксировать молек.азот. |
- |
|
Дыхание(аэробное и анаэробное), хемосинтез и |
Дыхание, фотосинтез у раст., питание(аэро- и |
|
фотосинтез |
анаэробы, автотрофы-хемо и фото, гетеротрофы) |
4. Принцип компартментации. Биологическая мембрана.
Высокая упорядоченность внутреннего содержимого клетки достигается путём компартментации её объёма – подразделения на отсеки, отличающиеся деталями хим.состава. Компартментация – пространственное разделение веществ и процессов в клетке. Компартменты – отсеки, ячейки – ядро, митохондрия, пластиды, лизосомы, вакуоли, т.к. образ мембраны.
Рис. 2.3. Компартментация объема клетки с помощью мембран:
1—ядро, 2—шероховатая цитоплазматическая есть, 3—митохондрия, 4—транспортный цитоплазматический пузырек, 5—лизосома, 6— пластинчатый комплекс, 7 — гранула секрета
Билипидный слой – гидрофобные хвосты – внутрь, гидрофильные головки – наружу. Мембранные белки:
периферические (примыкают к билипидному слою) – связ с липидными головками с помощью ионных связей; легко экстрагируются из мембран.
интегральные белки (пронизывающие – имеют каналы-поры, через к-рые проходят водорастворимые в-ва; погруженные белки (полуинтегральные) – пронизывают наполовину) – взаимодействуют с липидами на основе гидрофобных связей.
Мембранные липиды:
фосфолипиды – ост-к ж.к. – идеальный компонент для реализации барьерной ф-ции
гликолипиды – ост-к ж.к. + ост-к а/к
холестерол – стероидный липид, ограничив подвижн-ть липидов, уменьшает текучесть, стабилизирует мембрану.
Ф-ции мембраны: барьерная (защищает внутр содерж-е клетки), поддерживает постоянную форму кл-ки; обеспечивает связь клеток; пропускает внутрь кл-ки необходимые в-ва (избират прониц-ть – мол-лы и ионы проходят через мембрану с различной скоростью, чем больше размер, тем меньше скор-ть).
Свойства мембраны:
-билипидный слой способен к самосборке;
-увелич-е пов-ти мембраны за счёт встраивания в неёмембранных пузырьков (везикул);
-белки и липиды ассиметрично расположены в плоскости мембраны;
-белки и липиды могут перемещаться в плоскости мембраны в пределах слоя
(латеральное перемещ-е);
-наружн и внутр пов-ти мембраны имеют разный заряд.
-мембрана обеспечивает разделение заряженных частиц и поддержание разности потенциалов
5. Принцип клеточной компартментации. Организация и свойства биологической мембраны. История изучения.
См. 4 вопрос.
История изучения:
1902, Овертон находит липиды в составе пзазматической мебраны.
1925, Гортер и Грендел показывают наличие двойного слоя липидов в мембране эритроцитов.
1935, «бутербродная» модель Даниелли и Давсона (липидный бислой между двумя слоями белков)
Накопл-е фактов, необъяснимых с позиции «бутербродной» мембраны (мембраны очень динамичны)
1962, Мюллер создает плоскую модель искусственной мембраны 1957-1963, Робертсон формулирует понятие элементарная биологическая мембрана.
1972, создание Зингером и Николсоном жидкостно-мозаичной модели мембраны.
6.Структурная организация и свойства биологических мембран.
См. 5 вопрос
7.Мембранные белки и липиды.
Мембранные белки:
периферические (примыкают к билипидному слою) – связ с липидными головками с помощью ионных связей; легко экстрагируются из мембран.
интегральные белки (пронизывающие – имеют каналы-поры, через к-рые проходят водорастворимые в-ва; погруженные белки (полуинтегральные) – пронизывают наполовину) – взаимодействуют с липидами на основе гидрофобных связей.
Мембранные липиды:
фосфолипиды – ост-к ж.к. – идеальный компонент для реализации барьерной ф-ции
гликолипиды – ост-к ж.к. + ост-к а/к
холестерол – стероидный липид, ограничив подвижн-ть липидов, уменьшает текучесть, стабилизирует мембрану.
8.Явление осмоса в растительных и животных клетках.

Энергия АТФ, непосредственно или будучи перенесена на другие макроэргические соединения (например, креатинфосфат), в разнообразных процессах преобразуется в тот или иной вид работы. Одна из них осмотическая (поддержание перепадов концентрации веществ)
Осмос - диффузия (передвижение мол-л по градиенту конц-ции - из обл выс конц в обл низк конц) воды через полупрониц мембраны.
Враст кл-ке: Плазмолиз (когда жарко) - отток воды, содерж-е кл-ки сжим-ся и отходит от клет стенки. Деплазмолиз (прохл-но+полить) - кл-ки набух и прижим к клет стенке, подчин тургорному давл-ю (тургор - внутр гидростатич давл, вызывающее натяж-е кл стенки). Клеточная стенка способна растягиваться до определенного предела, после чего оказывает сопротивление - вытеснение воды из клеток происходит с такой же скоростью, с которой она в них поступает. (! прочность клеточной стенки не даёт раст. клеткам, в отличие от животных, лопнуть под напором).
Вжив кл-ке: изотонич р-р - норма, гипертон р-р - сморщив-е, гипотонич р-р - набух-е, затем лопаются-лизис.
Рис. 1. Осмос в искусственной системе. Трубку, содержащую раствор глюкозы и закрытую с одного конца мембраной,пропускающей воду, но не пропускающей глюкозу, опускают закрытым концом в сосуд с водой. Вода может проходить через мембрану в том и в другом направлении; однако молекулы глюкозы в трубке мешают движению соседних молекул воды, и потому больше воды входит в трубку, чем выходит из нее. Раствор поднимается в трубке до тех пор, пока давление его столба не станет достаточным для того, чтобы вытеснять воду из трубки с такой же скоростью, с какой она поступает внутрь.
Осмос - процесс одностороннего проникновения молекул растворителя через полунепроницаемую мембрану в сторону большей конц. растворенного вещ-ва. отчего зависит осмос? во-первых, от общей концентрации всех растворенных частиц по обе стороны от мембраны, ну а во-вторых от давления, создаваемого каждым "р-ром" (понятие осмотического давления: такое давление на раствор, обусловленное стремлением системы (ну т.е. клетки) выровнить конц. р-ра в обеих средазх, разделенных мембраной). Наличие воды необходимо для норм. протекания всех ппроцессов, и именно благодаря осмосу происходит "обводнение" клеток и структур. У КЛЕТОК НЕТ СПЕЦ, МЕХАНИЗМА ДЛЯ НАСАСЫВАНИЯ И ОТКАЧИВАНИЯ ВОДЫ НЕПОСРЕДСТВЕННО! - поэтому приток и отток воды регулируется изменением конц. вещ-в. Клеточная стенка способна растягиваться до определенного предела, после чего оказывает сопротивление - вытеснение воды из клеток происходит с такой же скоростью, с которой она в них поступает. (! прочность клеточной стенки не даёт раст. клеткам, в отличие от животных, лопнуть под напором).
9. Особенности строения растительных клеток. Осмотические свойства растительных клеток.
Особ строен растит кл-к: жёстк целлюлозопектинов стенка,пластиды, вакуоли с клет соком.
Жёсткость кл стенки предотвращ от чрезмерн набух-я и разрыва, обуславл потерю спос-ти к передвиж-ю. За счёт роста вакуоли увел размер кл-ки, игр важн роль в регуляции поступл-я воды в кл-ку,содерж растит антибиотики,к-ые убив микроорг-мы и микроскопич грибы. Пластиды-неоднородн группа органелл растит кл-ки (хлоропл-ты, хромопл-ты и лейкопласты)
Фотос-з –синт-з сложн орг в-в из неорг приучаст солн света Свет фаза 1-поглощ-е света хлорофиллом,возбужд-е его е.2-возбужд е перемещ по цепи переноса,отдавая лишн энерг на синтез АТФ 3-фотолиз воды (итог-синтез АТФ+фотолиз воды с выдел О2) Темн фаза 1-улавлив-ся СО2 2-синтез глюкозы из СО2 с пом энерг АТФ
Отличие растительной от животной клетки:
ВАКУОЛЬ. Окруж.мембраной-стонопласт.Компартмент, связанный с неподвижн обр жизни растит кл+ПЛАСТИДЫ(хлоропласты, хромопласты, лейкопласты)
Функции:
Накопительная(вода, глю, к-ты, фруктоза)+ненужные и невыводимые вещ-ва Алкалоиды-биологич. Активные вещ-ва; Пигменты(окраска зависит от рН) Поддержание осмотического давления(Тургор) Защитная(Бактериологические св-ва-фитонциды)
Ферментативная (роль мезосомы)
Нет клеточн.центра! Не способна к фагоцитозу(мешает клет.стенка)!
Механич прочность клет.стенок позволяет сущ в гипотонической среде, где в кл ОСМОТИЧЕСКИМ путем поступает вода. По мере поступления воды в клетку, возникает давление, препятствующее дальнейшему пост воды. Избыточное гидростатическое давление в клетке-ТУРГОР-обеспечение роста, сохр формы растением, определ положение в пространстве, противостояние механич воздействиям.
10. Поверхностный аппарат клетки. Пассивный транспорт.
1. Плазматическая мембрана Билипидный слой – гидрофобные хвосты – внутрь, гидрофильные головки – наружу. Мембранные белки:
периферические (примыкают к билипидному слою) – связ с липидными головками с помощью ионных связей; легко экстрагируются из мембран.
интегральные белки (пронизывающие – имеют каналы-поры, через к-рые проходят водорастворимые в-ва; погруженные белки (полуинтегральные) – пронизывают наполовину) – взаимодействуют с липидами на основе гидрофобных связей.
Мембранные липиды:
фосфолипиды – ост-к ж.к. – идеальный компонент для реализации барьерной ф-ции
гликолипиды – ост-к ж.к. + ост-к а/к
холестерол – стероидный липид, ограничив подвижн-ть липидов, уменьшает текучесть, стабилизирует мембрану.
Ф-ции мембраны: барьерная (защищает внутр содерж-е клетки), поддерживает постоянную форму кл-ки; обеспечивает связь клеток; пропускает внутрь кл-ки
необходимые в-ва (избират прониц-ть – мол-лы и ионы проходят через мембрану с различной скоростью, чем больше размер, тем меньше скор-ть).
2.Надмембранный комплекс – гликокаликс
олигосахариды гликолипидов и гликопротеинов,
периферич белки,
надмембр части интегр белков.
Ф-ции: изолирующая, рецепторная, маркировочная, ферментативная 3. Субмембранный комплекс – примембранная часть цитоскелета с белками,обеспечивщими её связь с мембр)
Ф-ции: поддерж-е формы, изменение пов-ти (при фагоцитозе и пиноцитозе)
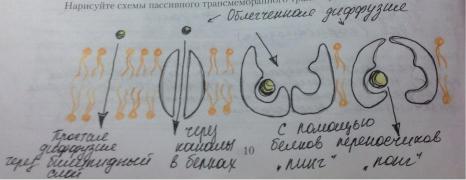
Пассивный транспорт-транспорт вещест, который осуществляется по градиенту концентрации и не требует затрат энергии. Если молекула заряжена, то на ее транспорт влияют как градиент концентрации, так и разница электрических потенциалов со стороны мембраны (мембранный потенциал). Вместе градиент концентрации и электрический градиент составляют электрохимический градиент. Активный и пассивный транспорт осуществляется с участием мембранных белков: белки переносчики (изменяют свою конформацию) и каналообразующие (формируют заполненные водой поры, пронизывающие липидный бислой; такие белковые каналы обладают ионной избирательностью, могут открываться при действии различных факторов, таких как: изменение мембранного потенциала, механическая стимуляция, при связывании с определенным лигандом.
Примеры пассивного транспорта:
Диффузия-передвижение молекул по градиенту концентрации, которое может происходить либо через билипидный слой (так транспортируются растворимые в липидах неполярные и слобополярные молекулы-кислород, углекислый газ, стероидные гормоны…), либо через поры, образуемые мембранными белками (так, например, перемещаются молекулы воды).
Облегченная диффузия-пассивный транспорт, в котором участвует транспортный белок (переносчик или каналообразующий). Белок-переносчик может пребывать в двух конформационных состояниях. В состоянии «пинг» этот белок соединяется с транспортируемой молекулой или ионом. Это вызывает его преход в состояние «понг», в котором он отдает транспортируемое вещество на противоположной стороне мембраны. Освободившись от транспортируемого вещества белок принимает исходное состояние (конформацию).
11. Поверхностный аппарат клетки. Активный транспорт.
См. 10 вопрос.
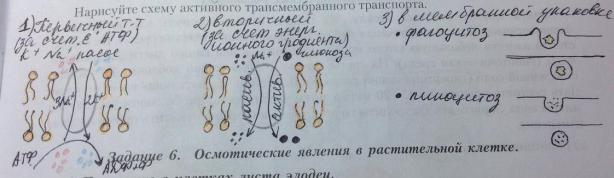
Активный транспорт-транспорт веществ, идущий с затратой энергии. В этом процессе могут участвовать как отдельные белки, так и обширные участки поверхностного аппарата клетки. Такой транспорт может осуществляться непосредственно за счет энергии АТФ (первичный активный транспорт), а также за счет энергии ионных градиентов (вторичный активный транспорт).
Пример первичного активного транспорта: калий-натриевый насос (ионы натрия-из клетки, ионы калия-в клетку, используя энергию АТФ). Фермент-калий-натриевая АТФаза, представляющая собой трансмембранный белок клеточной мембраны. Часть молекулы АТФ-азы, обращенная в сторону цитоплазмы, имеет центры связывания ионов натрия и калия и центр гидролиза АТФ. После присоединения трех ионов натрия АТФ-аза расщепляет АТФ с образованием АДФ и остатка фосфорной кислоты, котрый присоединяется к молекулы АТФ-азы (фосфорилирование). Фосфорилирование белка вызывает изменение его конформации: ионы натрия оказываются перенесенными на наружную сторону мембраны. Здесь происходит присоединение двух ионов калия и отщепление ионов натрия (с затратой энергии фосфорилирования). Остаток фосфорной кислоты отщепляется, и молекула белка принимает исходную конформацию, а ионы калия переносятся на внутреннюю сторону мембраны, где отщепляются. Таки образом поддерживается разность концентраций ионов в клетке и наружной среде.
Системы вторичного активного транспорта приводятся в действие за счет энергии, запасенной в ионных градиентах, а не путем прямого гидролиза АТФ. Они работают как котранспортные системы: одни-по принципу симпорта, другие-по принципу антипорта. В животных клетках котранспортным ионом обычно является натрий., который участвует в процессе всасывания глюкозы. Молекулы глюкозы и ионы натрия связываются с различными участками на белке-переносчике. Он меняет свою конформацию. Глюкоза и натрий оказываются на внутренней поверхности плазмалеммы. Ион натрия стремится выйти из клетки по своему электрохимическому градиенту, котрый обеспечивает активный транспорт глюкозы. Чем выше градиент, тем выше скорость всасывания глюкозы. Ионы натрия выкачиваются из клетке калий-натриевой АТФ-азой, которая косвенно контролирует транспорт глюкозы. Переход глюкозы в кровь происходит путем облегченной диффузии. Энергия ионный градиентов используется также для транспорта аминокислот через плазмалемму.
12. Поверхностный аппарат клетки. Транспорт макромолекул.
См. 10 вопрос
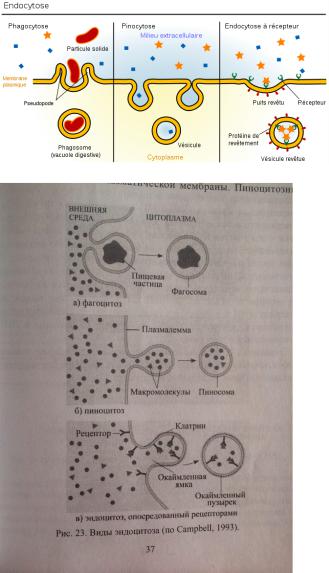
Эндоцитоз - это образование пузырьков путем выпячивания плазматической мембраны при поглощении твердых частиц-макромолекул (фагоцитоз) или растворенных веществ (пиноцитоз). Возникающие при этом гладкие или окаймленные эндоцитозные пузырьки называют также фагосомами или пиносомами.
Путем эндоцитоза осуществляются:
-питание (яйцеклетки поглощают таким способом желточные белки, фагосомами являются пищеварительные вакуоли простейших); -защитные и иммунные реакции (лейкоциты поглощают чужеродные частицы и иммуноглобулины);
-транспорт (почечные канальцы всасывают белки из первичной мочи). -поглощение ЛНП(липопротеинов низк плотн)
(Поглощение животными клетками холестерола из внеклеточной среды осуществляется за счет эндоцитоза, опосредуемого рецепторами. Основная часть холестерола переносится кровью в виде ЛПНП .
Молекулы ЛНП связываются со специальными рецепторами в плазмолемме клетки рецепторы, соединённые с лигандом(ЛНП) перемещаются по клеточной поверхности и накапливаются в окаймлённых ямках(их субмембранный аппарат содержит клатрин) из ямок обр-ся путём впячивания эндоцитозные пузырькипоступают в цитоплазму, теряют клатриновую оболочкусливаются с эндосомой, где лиганд отделяется от рецептораот мембраны эндосомы отделяются 2 вида пузырьков:одни содержат ЛНП и сливаются с
лизосомой, где из ЛНП выщепляется холестерол, другие содержат свободные рецепторы, которые возвращаются в мембрану.)
Избирательный эндоцитоз определенных веществ (желточных белков, иммуноглобулинов и т. п.) происходит при контакте этих веществ с субстрат-специфическими рецепторными участками на плазматической мембране.
Субстраты, попадающие в клетку путем эндоцитоза, расщепляются ("перевариваются"), накапливаются (например, желточные белки) или снова выводятся с противоположной стороны клетки путем экзоцитоза.
13.Поверхностный аппарат эукариотической клетки. Строение и функции.
См. 10 вопрос.
14.Транспортная роль белков плазмалеммы.
См.10 и 11 вопросы.
15.Активный транспорт веществ через мембрану. К+- Nа+ насос.
См. 11 вопрос
16.Поверхностный аппарат клетки. Транспорт макромолекул.
См. 12 (то же самое)
17. Рецепторная роль плазмалеммы.
Плазмалемма, биологическая мембрана, содержит различные сигнальные белки. Все водорастворимые и некоторые жирорастворимые сигнальные молекулы присоединяются к спецефическим белковым рецепторам на поверхности клеток-мишений. В зависимости от способа перечади сигнала внутрь клетки рецепторы можно разделить на три класса. 1. Каналообразующие рецепторы-регулируемые нейромедиаторами ионные каналы, участвующие в быстрой синаптической передачи импульсма. 2-каталитические рецепторыпри активации лигандом начинают работать как ферменты, фосфорилирую определенные белки.(напр, рецепторы инсулина) 3. Рецепторы, сопряженные с G- белками. Активация этих рецепторов запускает целую цепь реакций, изменяющих концентраци. В цитоплазме малых сигнальных молекул, которые называются вторичными посредниками. Наиболее важными вторичными посредниками являются циклические АМФ и Са. Эти молекулы могут влиять на поведение белков-мишеней в клетке, активирую обмен веществ.
Аденилатциклазная система:мембрана скелетных мыщц, жировые клетки, клетки сердцечной мыщцы. Сигнальными молекулами для этой системы служат многие гормоны и локальные химические медиаторы. Когда соответствующий лиганд ( например, адреналин) взаимодействует с рецептором, происходят конфармационные изменения рецепторного белка. Это вызывает структурную перестройку G-белка, находящегося в мембране. G-белок в этом состоянии активизирует адениатциклазу-фермент, также распологающийся в мембране. Активизированнная аденилатциклаща катализирует превращение АТФ в ц-АМФ. Ц-Амф воздействует на ферменты протеинкиназы, которые участвуют во многих метаболических процессах в клетке.