

BIO_-_vsyo
.pdf4. Механизмы сохранения нуклеотидной последовательности ДНК. Химическая стабильность. Репарация.
Репарация - исправл-е поврежд-ний в мол-ле ДНК.
Фотореактивация (светов репарация)-этот вид репарации позвол устранять димеры,возник между двумя соседними пиримидин-ми основ-ями под дей-ем УФ-излуч-я. Происх под дей-ем фотолиазы, к-рый активир квантами видимого света. Ферм-т соедин с поврежд-й ДНК, разъедин возникшие в димерах связи и восстанавлив водор связи между комплемент нуклеотидами.
Эксцизионная репарация (темнов репарация). Эксцизия-вырез-е. Устран поврежд-я, появившиеся под влиянием ионизирующей радиации, хим в-в. Поврежд уч-к нити вырезся, и синтезир-ся норм посл-ть нуклеотидов. Этапы: 1-ферм-т эндонуклеаза «узнаёт» поврежд уч-к ДНК; 2-эндонуклеаза разрезает одну нить мол-лы ДНК вблизи поврежд-я; 3- экзонуклеаза вырез поврежд уч-книти ДНК; 4-ДНК-полимераза провод матричн синтез нов цепи; 5-лигаза соедин новообразов уч-к с нитью ДНК.

Пострепликативн репарация часть поврежд-й ДНК не успев репариров-ся путём фотореактивации или эксцизии до репликации. При репликации поврежд уч-к не может использ в кач-ве матрицы. Репликация вынужд пропуст этот уч-к, оставляя брешь в дочерней нити.Т.о. образующ-ся дочерн мол-лы ДНК различны по природе. Одна из них содерж неповрежд родит цепь и норм дочерн комплемент-ую цепь. Др имеет родит цепь, несущую поврежд-е, и дочерн цепь с брешью. Пострепликативн репарация происх путём рекомбинации между дочерней, имеющей брешь, нитью и соотв-щей ей норм-й матер-й нитью др мол-лы. Образовавш-ся в матер-й цепи пробел может быть застроен обычным путём репаративного синтеза на комплемент-ой ей норм-й дочерней цепи. Поврежд уч-к может быть исправлен путём фотореактивации или эксцизионной репарации.
5. Способ записи генетической информации в молекуле ДНК. Биологический код и его свойства.
Генетич код - способ записи инф-ции об аминок-тах белка при помощи нуклеотидов ДНК.
Свойства:
1-триплетность (одна а/к кодируетсяся тремя нуклеотидамими, 3 нуклеотида-триплет) 2-избыточность (нек-рые а/к кодируются несколькими триплетами)
3-однозначность (каждому триплету соответствует одна а/к) 4-универсальность (для всех орг-в на Земле генетический код одинаков) 5-линейность (читается последовательноно)
6. Уникальные свойства ДНК: самоудвоение, самовосстановление структур.
Смотри 3 и 4 вопросы
7. Матричный синтез как специфическое свойство живого.
Матричный синтез 3 типа:
Синтез ДНК - репликация - самоудв-е мол-л ДНК,к-ое обычно происх перед дел-ем кл-ки. Во время репликацииматер мол-ла раскручив, и комплемент нити её разъедин(образ репликативн вилка) Формир-е репликат вилки происх под дей-ем ферментов геликазы и топоизомеразы. Геликаза разрыв водор связи между комплемент-ными нуклеотидами и разъедин нити, топоизомераза сним напряж-е, возникающее при этом в мол-ле. Одиночн нити матер мол-лы служат матрицами для синтеза дочерних комплемент-х нитей. С одиночн нитями связыв SSB-белки(дестабилизирующие белки),к-ые не дают им соедин в двойн спираль. В рез-те репликации образ две одинак мол-лы ДНК,полностью повторяющие матер мол-лу. При этом кажд нов мол-ла сост из одной нов и одной стар цепи. Комплемент нити мол-лы ДНК антипараллельны. Наращив-е полинуклеотидной цепи всегда происх в направл от 5' конца к 3' концу. Вследствие этого одна нить лидирующ (3' конец в основании репликативной вилки), а др-запаздывающ (5' конец в основ вилки) и поэтому строится из фрагменьов Оказаки, растущих от 5' к 3' концу. Фрагменты Оказаки – это участки ДНК, которые у эукариот имеют длину 100-200 нуклеотидов, у прокариот – 1000-2000 нуклеотидов.
Синтез цепи ДНК осуществляет фермент ДНК-полимераза. Она наращив дочерн цепь, присоединяя к её 3' концу нуклеотиды, комплементарные нуклеотидам материнской цепи. Особ-ть ДНК-полимеразы сост в том, что она не может начать работу на «пустом месте», не имея 3' конца дочерней нити. Поэтому синтез лидирующей нити и синтез каждого фрагмента Оказаки начинает фермент праймаза. Это разновидность РНК-полимеразы. Праймаза способна начать синтез новой полинуклеотидной цепи с соедин-я двух нуклеотидов. Праймаза синтезирует из РНК-нуклеотидов короткие затравки - праймеры. Их длина около 10 нуклеотидов. К 3' концу праймера ДНК полимераза начин присоединять ДНК-нуклеотиды.
Фермент экзонуклеаза удал праймеры. ДНК-полимераза достраивает фрагменты Оказаки, фермент лигаза сшивает их.
Синтез РНК - транскрипция - синтез РНК на матрице ДНК (у эукариот в ядре, у прокариот-в цитоплазме). В процессе транскрипции строится комплемент копия одной из нитей ДНК. В рез-те транскрипции синтезир-ся иРНК, рРНК и тРНК. Транскр-ю осущ РНК-полимераза. У эукариот транскрипцию оскществл три разные РНК-полимеразы:
-РНК-полимераза I синтезир рРНК
-РНК-полимераза II синтезир иРНК
-РНК-полимераза III синтезир тРНК
РНК-полимераза связыв-ся с молекулой ДНК в области промотора. Промотор – это участок ДНК, отмечающий начало транскрипции. Он расположен перед структурным геном. Присоединившись к промотору, РНК-полимераза раскручивает участок двойной спирали ДНК и раздел комплемент-ые цепи. Одна из двух цепей – смысловая – служит матрицей для синтеза РНК. Нуклеотиды РНК комплементарны нуклеотидам смысловой цепи ДНК. Транскрипция идёт от 5' конца к её 3' концу. РНК-полимераза отдел синтезиров-ый уч-к РНК от матрицы и восстанавливает двойную спираль ДНК.

Транскрипция продолжается до тех пор, пока РНК-полимераза не доёдет до терминатора. Терминатор – это уч-к ДНК, обозначающий конец транскрипции. Достигнув терминатора, РНК-полимераза отделяется и от матричной ДНК и от новосинтезированной молекулы РНК.
Транскр-я дел на 3 этапа:
Инициация – присоед-е РНК-полимеразы и помогающих ей белков-факторов транскрипции к ДНК и начало их работы.
Элонгация-наращив- полинуклеот-ой цепи РНК. Терминация-оконч-е синтеза мол-лы РНК.
Синтез белка - трансляция - процесс синтеза полипепт-ной цепи, проходящей на рибосоме. Происх в цитоплазме. Рибосома сост из двух субъединиц: большой и малой. Субъединицы построены из рРНК и белков. Неакт рибосома находится в цитоплазме в диссоциированном виде. Активная рибосома собирается из двух субъединиц, приэтом в ней образ-ся активные центры, в том числе – аминоацильный и пептидильный. В аминоацильном центре происход образ-е пептидной связи. Транспортные РНК специфичны, т.е. одна тРНК может перенос только одну определ-ую а/к. Эта а/к зашифрована кодоном, которому комплементарен антикодон тРНК. В процессе трансляции рибосома переводит последоват-ть нуклеотидов иРНК в последоват-ть а/к полипептидной цепи.
Трансляция дел на 3 этапа.
Инициация-сборка рибосомы на инициирующем кодоне иРНК и начало её работы. Инициация начинается с того, что с иРНК соедин-ся малая субъединица рибосомы и тРНК, несущая метионин, к-рый соответствует инициирующему кодону АУГ. Затем к этому комплексу присоедин-ся большая субъединица. В рез-те инициирующий кодон оказыв-ся в пептидильном центре рибосомы, а в аминоацильном центре наход-ся первый значащий кодон. К нему подходят различные тРНК, а останется в рибосоме только та, антикодон к-рой комплементарен кодону. Между комплемент-ми нуклеотидами кодона и антикодона образ-ся водородные связи. В итоге в рибосоме с иРНК оказыв-ся временно связаны две тРНК. Кажд тРНК принесла в рибосому а/к, зашифрованную кодоном иРНК. Между этими а/к образ-ся пептидная связь. После этого тРНК, принесшая метионин, отдел-ся от своей а/к и от иРНК и уходит из рибосомы. Рибосома перемещ-ся на один триплет от 5' конца к 3' концу иРНК.
Элонгация – процесс наращив-я полип-ой цепочки. В аминоацильный центр рибосомы будут подходить различн тРНК. Процесс узнавания тРНК и поцесс формирования пептидной связи будет повтор-ся до тех пор, пока в аминоацильном центре рибосомы не окажется стоп-кодон.
Терминация – заверш-е синтеза полипептида и диссоциация рибосомы на две субъединицы. Существ три стоп-кодона: УАА, УАГ и УГА. Когда один из них оказыв-ся в аминоацильном центре рибосомы, с ним связыв-ся белок – фактор терминации трансляции. Это вызывает распад всего комплекса.
8. РНК. Виды РНК и их биологическая роль.
РНК (рибонуклеиновая к-та)- полимер, мономерами к-го явл нуклеотиды. Нуклеотиды РНК сост из пятиугл-го сахара рибозы,ост-ка фосф-й к-ты и азотистого основ-я-цитозина, гуанина,аденина или урацила. Нуклеотиды соед между собой при помощи фосфодиэфирной ковал-й связи.
По выполн-м ф-циям РНК делят на:
тРНК сост из 80-100 нуклеотидов, содерж в цитоплазме. Ф-ция: перенос а/к к месту синтеза белка (в рибосомы). Полинуклеотидн цепь тРНК образ три петли.

рРНК сост из 3-5 тыс нуклеотидов, составл-х основу рибосомы.
иРНК от 300 до 30000 нуклеотидов в завис-ти от длины уч-ка ДНК, на котором синтезируется иРНК, содерж в ядре и цитоплазме. Ф-ция: перенос инф-ции о стр-ре белка от ДНК к месту синтеза белка в рибосомах
9. Роль РНК в реализации наследственной информации. Синтез белка.
Синтез белка осущ-ся в 2 этапа
Транскрипция-синтез РНК на матрице ДНК (у эукариот в ядре, у прокариот-в цитоплазме). В процессе транскрипции строится комплемент копия одной из нитей ДНК. В рез-те транскрипции синтезир-ся иРНК, рРНК и тРНК. Транскр-ю осущ РНКполимераза. У эукариот транскрипцию оскществл три разные РНК-полимеразы:
-РНК-полимераза I синтезир рРНК
-РНК-полимераза II синтезир иРНК
-РНК-полимераза III синтезир тРНК
РНК-полимераза связыв-ся с молекулой ДНК в области промотора. Промотор – это участок ДНК, отмечающий начало транскрипции. Он расположен перед структурным геном. Присоединившись к промотору, РНК-полимераза раскручивает участок двойной спирали ДНК и раздел комплемент-ые цепи. Одна из двух цепей – смысловая – служит матрицей для синтеза РНК. Нуклеотиды РНК комплементарны нуклеотидам смысловой цепи ДНК. Транскрипция идёт от 5' конца к её 3' концу. РНК-полимераза отдел синтезиров-ый уч-к РНК от матрицы и восстанавливает двойную спираль ДНК. Транскрипция продолжается до тех пор, пока РНК-полимераза не доёдет до терминатора. Терминатор – это уч-к ДНК, обозначающий конец транскрипции. Достигнув терминатора, РНК-полимераза отделяется и от матричной ДНК и от новосинтезированной молекулы РНК.
Транскр-я дел на 3 этапа:
Инициация – присоед-е РНК-полимеразы и помогающих ей белков-факторов транскрипции к ДНК и начало их работы.
Элонгация-наращив- полинуклеот-ой цепи РНК. Терминация-оконч-е синтеза мол-лы РНК.
Трансляция - процесс синтеза полипепт-ной цепи, проходящей на рибосоме. Происх в цитоплазме. Рибосома сост из двух субъединиц: большой и малой. Субъединицы построены из рРНК и белков. Неакт рибосома находится в цитоплазме в диссоциированном виде. Активная рибосома собирается из двух субъединиц, приэтом в ней образ-ся активные центры, в том числе – аминоацильный и пептидильный. В аминоацильном центре происход образ-е пептидной связи. Транспортные РНК специфичны, т.е. одна тРНК может перенос только одну определ-ую а/к. Эта а/к зашифрована кодоном, которому комплементарен антикодон тРНК. В процессе трансляции рибосома переводит последоват-ть нуклеотидов иРНК в последоват-ть а/к полипептидной цепи.
Трансляция дел на 3 этапа. Инициация-сборка рибосомы на инициирующем кодоне иРНК и начало её работы. Инициация начинается с того, что с иРНК соедин-ся малая субъединица рибосомы и тРНК, несущая метионин, к-рый соответствует инициирующему кодону АУГ. Затем к этому комплексу присоедин-ся большая субъединица. В рез-те инициирующий кодон оказыв-ся в пептидильном центре рибосомы, а в аминоацильном центре наход-ся первый значащий кодон. К нему подходят различные тРНК, а останется в

рибосоме только та, антикодон к-рой комплементарен кодону. Между комплемент-ми нуклеотидами кодона и антикодона образ-ся водородные связи. В итоге в рибосоме с иРНК оказыв-ся временно связаны две тРНК. Кажд тРНК принесла в рибосому а/к, зашифрованную кодоном иРНК. Между этими а/к образ-ся пептидная связь. После этого тРНК, принесшая метионин, отдел-ся от своей а/к и от иРНК и уходит из рибосомы. Рибосома перемещ-ся на один триплет от 5' конца к 3' концу иРНК.
Элонгация – процесс наращив-я полип-ой цепочки. В аминоацильный центр рибосомы будут подходить различн тРНК. Процесс узнавания тРНК и поцесс формирования пептидной связи будет повтор-ся до тех пор, пока в аминоацильном центре рибосомы не окажется стоп-кодон.
Терминация – заверш-е синтеза полипептида и диссоциация рибосомы на две субъединицы. Существ три стоп-кодона: УАА, УАГ и УГА. Когда один из них оказыв-ся в аминоацильном центре рибосомы, с ним связыв-ся белок – фактор терминации трансляции. Это вызывает распад всего комплекса.
10. “Центральная догма” молекулярной биологии. Понятие об обратной транскрипции. Современные проблемы генной инженерии.
«Центральная догма» биологии - это поток инф-ции от ДНК через РНК на белок. Характерен для всех живых орг-в, за исключ-ем нек-ых РНК-содержащих вирусов. Матричн природа синтеза нуклеин-х к-т и белков обеспеч высок точность воспроизвед-я инф-ции. Направл-е потока инф-ции от ДНК к стр-ре белка включ 3 типа матричных синтезов: синтез ДНК-репликация - самоудв-е мол-л ДНК,к-ое обычно происх перед делем кл-ки. Во время репликацииматер мол-ла раскручив, и комплемент нити её разъедин(образ репликативн вилка) Формир-е репликат вилки происх под дей-ем ферментов геликазы и топоизомеразы. Геликаза разрыв водор связи между комплементными нуклеотидами и разъедин нити, топоизомераза сним напряж-е, возникающее при этом в мол-ле. Одиночн нити матер мол-лы служат матрицами для синтеза дочерних комплемент-х нитей. С одиночн нитями связыв SSB-белки(дестабилизирующие белки),к- ые не дают им соедин в двойн спираль. В рез-те репликации образ две одинак мол-лы ДНК,полностью повторяющие матер мол-лу. При этом кажд нов мол-ла сост из одной нов и одной стар цепи. Комплемент нити мол-лы ДНК антипараллельны. Наращив-е полинуклеотидной цепи всегда происх в направл от 5' конца к 3' концу. Вследствие этого одна нить лидирующ (3' конец в основании репликативной вилки),а др-запаздывающ (5' конец в основ вилки) синтез РНК-транскрипция -синтез РНК на матрице ДНК (у эукариот в ядре,у прокариот-в цитоплазме). В процессе транскрипции строится комплемент копия одной из нитей ДНК. В рез-те транскрипции синтезир-ся иРНК,рРНК и тРНК. Транскр-ю осущ РНК-полимераза. Транскр-я дел на 3 этапа. Инициация – присоед-е РНК-полимеразы и помогающих ей белков-факторов транскрипции к ДНК и начало их работы. Элонгация-наращив- полинуклеот-ой цепи РНК. Терминация- оконч-е синтеза мол-лы РНК. синтез белка -трансляция - процесс синтеза полипепт-ной цепи, проходящей на рибосоме. Происх в цитоплазме. Дел на 3 этапа. Инициация-сборка рибосомы на инициирующем кодоне иРНК и начало её работы. Элонгация –процесс наращив-я полип-ой цепочки. Терминация –заверш-е синтеза полипептида и диссоциация рибосомы на две субъединицы. Обратн транскрипция - синтез комплементарной ДНК, к-ая может включ-ся в геном высших организмов. Например, матрица РНКразмножающегося вируса.
Современные проблемы генной инженерии Генная инженерия - это раздел молекул-й
биологии, прикладная молекул генетика, задачей к-й явл целенаправл-е конструиров-е новых, не существующих в природе сочетаний генов при помощи генетич-х и биохимич-х методов. Этич и религиозн аспекты.

11. Синтез белка в клетке. Генетический код. Функция информационной, транспортной и рибосомной РНК.
Синтез белка осущ-ся в 2 этапа
Транскрипция-синтез РНК на матрице ДНК (у эукариот в ядре, у прокариот-в цитоплазме). В процессе транскрипции строится комплемент копия одной из нитей ДНК. В рез-те транскрипции синтезир-ся иРНК, рРНК и тРНК. Транскр-ю осущ РНКполимераза. У эукариот транскрипцию оскществл три разные РНК-полимеразы:
-РНК-полимераза I синтезир рРНК
-РНК-полимераза II синтезир иРНК
-РНК-полимераза III синтезир тРНК
РНК-полимераза связыв-ся с молекулой ДНК в области промотора. Промотор – это участок ДНК, отмечающий начало транскрипции. Он расположен перед структурным геном. Присоединившись к промотору, РНК-полимераза раскручивает участок двойной спирали ДНК и раздел комплемент-ые цепи. Одна из двух цепей – смысловая – служит матрицей для синтеза РНК. Нуклеотиды РНК комплементарны нуклеотидам смысловой цепи ДНК. Транскрипция идёт от 5' конца к её 3' концу. РНК-полимераза отдел синтезиров-ый уч-к РНК от матрицы и восстанавливает двойную спираль ДНК. Транскрипция продолжается до тех пор, пока РНК-полимераза не доёдет до терминатора. Терминатор – это уч-к ДНК, обозначающий конец транскрипции. Достигнув терминатора, РНК-полимераза отделяется и от матричной ДНК и от новосинтезированной молекулы РНК.
Транскр-я дел на 3 этапа:
Инициация – присоед-е РНК-полимеразы и помогающих ей белков-факторов транскрипции к ДНК и начало их работы.
Элонгация-наращив- полинуклеот-ой цепи РНК. Терминация-оконч-е синтеза мол-лы РНК.
Трансляция - процесс синтеза полипепт-ной цепи, проходящей на рибосоме. Происх в цитоплазме. Рибосома сост из двух субъединиц: большой и малой. Субъединицы построены из рРНК и белков. Неакт рибосома находится в цитоплазме в диссоциированном виде. Активная рибосома собирается из двух субъединиц, приэтом в ней образ-ся активные центры, в том числе – аминоацильный и пептидильный. В аминоацильном центре происход образ-е пептидной связи. Транспортные РНК специфичны, т.е. одна тРНК может перенос только одну определ-ую а/к. Эта а/к зашифрована кодоном, которому комплементарен антикодон тРНК. В процессе трансляции рибосома переводит последоват-ть нуклеотидов иРНК в последоват-ть а/к полипептидной цепи.
Трансляция дел на 3 этапа. Инициация-сборка рибосомы на инициирующем кодоне иРНК и начало её работы. Инициация начинается с того, что с иРНК соедин-ся малая субъединица рибосомы и тРНК, несущая метионин, к-рый соответствует инициирующему кодону АУГ. Затем к этому комплексу присоедин-ся большая субъединица. В рез-те инициирующий кодон оказыв-ся в пептидильном центре рибосомы, а в аминоацильном центре наход-ся первый значащий кодон. К нему подходят различные тРНК, а останется в рибосоме только та, антикодон к-рой комплементарен кодону. Между комплемент-ми нуклеотидами кодона и антикодона образ-ся водородные связи. В итоге в рибосоме с иРНК оказыв-ся временно связаны две тРНК. Кажд тРНК принесла в рибосому а/к, зашифрованную кодоном иРНК. Между этими а/к образ-ся пептидная связь. После этого тРНК, принесшая метионин, отдел-ся от своей а/к и от иРНК и уходит из рибосомы. Рибосома перемещ-ся на один триплет от 5' конца к 3' концу иРНК.
Элонгация – процесс наращив-я полип-ой цепочки. В аминоацильный центр рибосомы будут подходить различн тРНК. Процесс узнавания тРНК и поцесс формирования пептидной связи будет повтор-ся до тех пор, пока в аминоацильном центре рибосомы не окажется стоп-кодон.
Терминация – заверш-е синтеза полипептида и диссоциация рибосомы на две субъединицы. Существ три стоп-кодона: УАА, УАГ и УГА. Когда один из них оказыв-ся в аминоацильном центре рибосомы, с ним связыв-ся белок – фактор терминации трансляции. Это вызывает распад всего комплекса.
Генетич код-способ записи инф-ции об аминок-тах белка при помощи нуклеотидов ДНК. Св-ва: 1-триплетность (одна а/к кодир-ся тремя нуклеот-ми, 3 нуклеотида-триплет) 2- избыточность(нек-рые а/к кодир-ся неск-ми триплетами) 3-однозначность(кажд триплету соотв одна а/к) 4-универсальность(для всех орг-в на Земле генетт код одинаков) 5- линейность (читается последоват-но)
По выполн-м ф-циям РНК делят на: тРНК сост из 80-100 нуклеотидов, содерж в цитоплазме. Ф-ция: перенос а/к к месту синтеза белка (в рибосомы). Полинуклеотидн цепь тРНК образ три петли. рРНК сост из 3-5 тыс нуклеотидов, составл-х основу рибосомы. иРНК от 300 до 30000 нуклеотидов в завис-ти от длины уч-ка ДНК, на котором синтезируется иРНК, содерж в ядре и цитоплазме. Ф-ция: перенос инф-ции о стр-ре белка от ДНК к месту синтеза белка в рибосомах
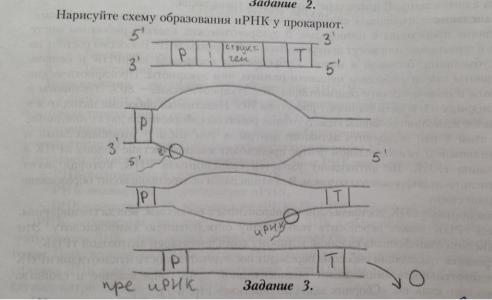
12. Особенности образования иРНК в клетках эу- и прокариот.
У прокариот новосинтезир-ые мол=лы иРНК на 5' конце им неинформативн уч-к, за к- рым следует инициирующий кодон АУГ, обозначающ начало трансляции. Далее наход информативн уч-к, на к-ром записана инф-ция о стр-ре белка. За ним следует стоп-кодон (УАА//УАГ//УГА), определ конец трансляции . На 3' конце наход концев неинформативн последоват-ть.
У эукариот новосинтезиров иРНК претерпев посттранскрипц измен-я-процессинг. К 5' концу пре-иРНК присоедин кэп-метилированный гуанозинтрифосфат. К 3' концу присоедин фрагмент, состоящ из 100-300 адениловых нуклеотидов –поли-А последоватть. Кэпирование(присоед-е кэп) и полиаденилиров-е (присоед-е поли-А последоват-ти) происх в ядре в момент оконч-я синтеза иРНК. В рез-те образ-ся пре-иРНК, к-ая им следующее строение: на 5' конце наход кэп, за ним неинформативная последоват-ть,
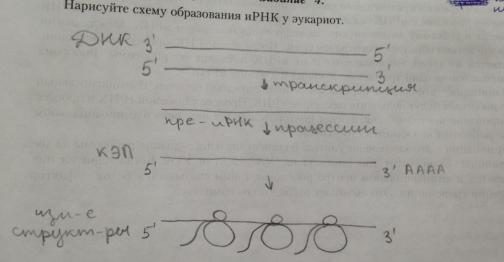
инициирующий кодон АУГ, далее – отрезок, где информативные уч-ки – экзоны – чередся с неинформативными – интронами. Этот отрезок заканчив стоп-кодоном, за к-рым идёт неинформативная последоват-ть и поли-А последоват-ть. У эукариот в ядре происх сплайсинг – вырез-е интронов и сшивание экзонов.
13. Прерывистая (экзонно-интронная) структура гена у эукариот. Сплайсинг. Альтернативный сплайсинг.
Прерывистая (экзонно-интронная) стр-ра гена у эукариот В конце 70-х годов было выяснено, что у эукариот имеются гены, которые содержат «лишнюю» ДНК, не представленную в молекуле иРНК. Они получили название мозаичных, прерывистых генов; генов, имеющих экзон-интронное строение. Первый продукт транскрипции – гетерогенная РНК – имеет экзонно-интронную прерывистую структуру. Вторичный экзоный продукт транскрипции – зрелая иРНК Сплайсинг - вырез-е из первичного транскрипта интронов (неинформативных уч-в) и сшивание экзонов (информативных уч-в).
Один ген может служить матрицей для неск-х различных белков, если происход альтернативный сплайсинг, т.е. в кач-ве интронов вырез-ся разн уч-ки мол-лы преиРНК. Альтернативный сплайсинг – из одного первого РНК-транскрипта в разных тканях образуется несколько разных по длине зрелых иРНК. Полипептиды также будут различны.
14. Экспрессия генетической информации у эукариот.
Экспрессия генет инф-ции у эукариот
У эукариот новосинтезиров иРНК претерпев посттранскрипц измен-я-процессинг. К 5' концу пре-иРНК присоедин кэп-метилированный гуанозинтрифосфат. К 3' концу присоедин фрагмент, состоящ из 100-300 адениловых нуклеотидов –поли-А последоватть. Кэпирование(присоед-е кэп) и полиаденилиров-е (присоед-е поли-А последоват-ти) происх в ядре в момент оконч-я синтеза иРНК. В рез-те образ-ся пре-иРНК, к-ая им следующее строение: на 5' конце наход кэп, за ним неинформативная последоват-ть, инициирующий кодон АУГ, далее – отрезок, где информативные уч-ки – экзоны – чередся с неинформативными – интронами. Этот отрезок заканчив стоп-кодоном, за к-рым идёт неинформативная последоват-ть и поли-А последоват-ть. У эукариот в ядре происх сплайсинг – вырез-е интронов и сшивание экзонов.
15. Экспрессия генетической информации у прокариот.
Экспрессия генетической информации у прокариот
У прокариот новосинтезир-ые мол=лы иРНК на 5' конце им неинформативн уч-к, за к- рым следует инициирующий кодон АУГ, обозначающ начало трансляции. Далее наход информативн уч-к, на к-ром записана инф-ция о стр-ре белка. За ним следует стоп-кодон (УАА//УАГ//УГА), определ конец трансляции . На 3' конце наход концев неинформативн последоват-ть.
16. Регуляция экспрессии генов у эукариот (на уровне транскрипции, процессинга и посттранскрипционном уровне).
Регул экспрессии генов у эукариот на ур-не транскрипции: регул-я за счёт конденсации и деконденсации ДНК. хр-мы типа ламповых щёток, инактивация х-хромосомы. Транскрипция происх только с эухроматина-деконденсированных уч-в ДНК. Поэтому переход гетерохроматина в эухроматин делает возможной транскрипцию, а обр переход останавливает её. Регул-я при участии энхансеров, промоторов и регуляторных белков:
Промоторы и энхансеры – спец регуляторн последоват-ти ДНК,к-ые участв в регул транскрипции у эукариот. Промотор-это уч-к ДНК длиной 100 нуклеотидов, располож-й перед геном. Энхансер-значительно удал от промотора уч-к ДНК,к-ый может быть расположен как перед структ геном, так и после него. С промоторами и энхансерами связ белки-регуляторы: активаторы-вызыв транскрипцию и репрессоры-подавл её. Регул экспрессии генов у эукариот на ур-не процессинга: процессинг-совок-ть