

BIO_-_vsyo
.pdfS-периоде происходит и удвоение числа генов рРНК, поэтому их количество коррелирует с плоидностью клетки.
Однако существуют случаи, когда гены рРНК подвергаются избыточной репликации. При этом дополнительная репликация генов рРНК происходит в целях обеспечения продукции большого количества рибосом. В результате такого сверхсинтеза генов рРНК их копии могут становиться свободными, экстрахромосомными. Эти внехромосомные копии генов рРНК могут функционировать независимо, в результате чего возникает масса свободных дополнительных ядрышек, но уже не связанных структурно с ядрышкообразующими хромосомами. Это явление получило название амплификации генов рРНК.
56. Ядро – система хранения, воспроизведения и реализации генетический информации.
Клеточное ядро отделено от содержимого клетки оболочкой. Функции ядра состоят в хранении наследственного материала (ДНК), его воспроизводстве (репликация ДНК) с целью передачи в ряду клеточных поколений (митоз), а также в реализации наследственной информации в ходе биосинтеза белка в жизнедеятельности клетки (транскрипция, процессинг пре-РНК транскриптов). В нем образуются структурные элементы - большая и малая субъединицы - цитоплазматических органелл рибосом, на которых в цитоплазме происходит образование полипептидов (простых белков).
Вядре выделяют ядерную оболочку, ядерный матрикс, ядрышко, хромосомы (хроматин), ядерный сок.
Вядре сосредоточена большая часть ДНК эукариотической клетки - 90%. Она распределена между ядерными структурами – хромосомами. Морфология хромосом меняется по стадиям клеточного цикла. При вхождении клетки в митоз материал хромосом приобретает плотную упаковку (митотическая форма), а вне митоза - рыхлую (интерфазная форма). хроматин - вещество хромосомы, а его состав и плотность упаковки различаются по длине хромосомы в зависимости от стадии это го цикла.
Хромосомы во взаимодействии с внехромосомными механизмами обеспечивают:
•хранение генетической информации;
•использование этой информации для воспроизводства и поддержания клеточной организации и функций;
•регуляцию считывания (транскрипция) наследственной информации;
•удвоение (репликация, самокопирование) генетического материала материнских клеток перед клеточным делением;
•передачу этого материала дочерним клеткам в процессе митоза. Первую из этих функций хромосома выполняет в обеих структурных формах - митотической и интерфазной, следующие три функции - в интерфазной форме, последнюю - в митотической форме. Хромосомная организация наследственного материала эукариот создает условия для тонкой регуляции генетических функций, репаративных процессов, минимизирующих объем нарушений молекулярной структуры ДНК, а также для рекомбинации ДНК в ходе мейоза при образовании половых клеток (см. кроссинговер, комбинативная генотипическая изменчивость).
Рис. 2.8. Структура клеточного ядра (схема): 1 - ядерная оболочка (две мембраны - внешняя и внутренняя, и перинуклеарное пространство); 2 - ядерная пора; 3 - конденсированный хроматин; 4 - диффузный хроматин; 5 - ядрышко (гранулярный и фибриллярный компоненты, в центральных светрых зонах находится р-ДНК); 6 - интерхроматиновые гранулы (РНП); 7 - перихрома-тиновые гранулы (РНП); 8 - перихроматиновые фибриллы (РНП); 9 - кариоплазма, ядерный сок
57. Организация эу- и гетерохроматина. Структура и химия хроматина.
Вядре сосредоточена большая часть ДНК эукариотической клетки - 90%.в составе хромосом. Материал хромосом - совокупность глыбок, зерен и волоконец – хроматина.
Химический состав хроматина (хромосом) эукариотической клетки
Большая часть объема хромосом представлена ДНК и белками. Заметные химические компоненты хромосом - РНК и липиды. Среди белков (65% массы хромосом) выделяют гистоновые (60-80%) и не-гистоновые. Также присутствуют полисахариды, ионы металлов (Ca, Mg) и др. Особое место среди хромосомных белков принадлежит гистонам.
Всоставе нуклеогистонового комплекса ДНК менее доступна ферментам нуклеазам, вызывающим ее гидролиз (функция защиты). Гистоны выполняют структурную функцию, участвуя в процессе компактизации хроматина. Гистоновые белки представлены пятью видами (фракциями): Н1, Н2А, Н2В, Н3 и Н4.
Число ядерных негистоновых белков превышает несколько сотен. Они удерживают «открытую» конфигурацию хроматина, «разрешающую» доступ к биоинформации ДНК, то есть ее транскрипцию.
К категории «временных» относятся цитозольные белки-рецепторы (функциональнотранскрипционные факторы), захватывающие сигнальные молекулы, в комплексе с которыми они проникают в ядро и их активируют.
РНК хромосом представлена продуктами транскрипции, еще не покинувшими место синтеза, - непосредственный продукт транскрипции генов или пре-и(м)РНК, пре-рРНК, пре-тРНК транскрипты. Некоторые виды РНК «временного внутриядерного пребывания» создают условия для основного процесса, выполняя сигнальную функцию. Так, репликация ДНК требует для своего начала образуемой «на месте» РНК-затравки (РНКпраймер), которая по завершении процесса разрушается здесь же в ядре.
Взависимости от степени компактизации материал интерфазных хромосом представлен эухроматином и гетерохроматином . Эухроматин - низкая степень компактизации и неплотная «упаковка» хромосомного материала. Эухроматин представлен, в основном, ДНК с уникальными последовательностями нуклеотидов. Гены из эухроматизированного участка хромосомы, оказавшись в гетерохроматизированномучастке или рядом с ним, обычно инактивируются.
Гетерохроматин отличается высокой степенью компактизации, то есть плотной «упаковкой» материала хромосомы. Большая его часть представлена умеренно или
многократно повторяющимися нуклеотидными последовательностями ДНК. К первым относятся мультикопийные гены гистонов, рибосомных и транспортных РНК.
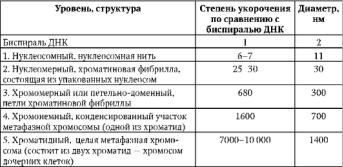
58. Уровни структурной организации хроматина. Компактизация хроматина.
На протяжении клеточного цикла хромосома сохраняет структурную целостность благодаря компактизации-декомпактизации (конденсация-деконденсация) хромосомного материала – хроматина. Вследствие компактизации при переходе хромосом из интерфазной формы в митотическую суммарный линейный показатель сокращается примерно в 7-10 тыс. раз.
Таблица 2.1. Последовательные уровни компактизации хроматина.
В образовании нуклеосомной нити ведущая роль принадлежит гистонам Н2А, Н2В, Н3 и Н4. Они образуют белковые тела или коры, состоящие из восьми молекул. Молекула ДНК комплексуется с белковыми корами, спирально накручиваясь на них-биспирали.
Свободную от контакта с корами ДНК наз линкерной(связующая). Отрезок ДНК + белк кор = нуклеосома. Благодаря нуклеосомам в промоторных участках ДНК заблокированы области инициации (начала) транскрипции. Для того чтобы инициаторный комплекс возник, нуклеосомы должны быть «вытеснены» из соответствующих фрагментов ДНК. Образование хроматиновой фибриллы диаметром 30 нм (второй уровень компактизации) происходит с участием гистона Н1, который, связываясь с линкерной ДНК, скручивает нуклеосомную нить в спираль.
На следующем петельно-доменном -укладка фибриллы диаметром 30 нм в петли. В этом процессе активная роль отводится негистоновым белкам. Основания петель «заякорены» в ядерном матриксе. Петля содержит от одного до нескольких генов(петельный домен).
На следующем уровне компактизации «сложенные» фибриллы превращаются в метафазные хроматиды (хромосомы будущих дочерних клеток).
Максимальная степень компактизации достигается на пятом уровне в структурах, известных как метафазные хромосомы с диаметром 1400 нм. Такая структура обеспечивает оптимальное решение задачи транспортировки генетического материала в дочерние клетки в анафазе митоза.
59. Динамика хромосомного материала в клеточном цикле.
|
|
СТЕПЕНЬ |
|
|
ДИАМЕТР |
УКОРОЧЕНИЯ |
|
СТРУКТУРА |
(относительно |
||
(в НМ) |
|||
|
БИСПИРАЛИ |
||
|
|
||
|
|
ДНК) |
|
* БИСПИРАЛЬ ДНК |
2 |
1 |
|
* НУКЛЕОСОМНАЯ НИТЬ |
11 |
7 |
|
* ХРОМАТИНОВАЯ ФИБРИЛЛА из |
|
|
|
УПАКОВАННЫХ (8-10) |
30 |
40 |
|
НУКЛЕОСОМ |
|
|
* ПЕТЛИ ХРОМАТИНОВОЙ |
|
|
|
ФИБРИЛЛЫ (ПЕТЕЛЬНО- |
300 |
1000 |
|
ДОМЕННАЯ |
|||
|
|
||
СТРУКТУРА) |
|
|
|
* ХРОМАТИДА – КОНДЕНСАЦИЯ |
|
|
|
МАТЕРИАЛА ХРОМОСОМ ПРИ |
700 |
1600 |
|
ВСТУПЛЕНИИ КЛЕТКИ В МИТОЗ |
|
|
|
* МЕТАФАЗНАЯ ХРОМОСОМА, |
|
|
|
МАКСИ-МАЛЬНАЯ СТЕПЕНЬ |
1400 |
8000 |
|
КОНДЕНСАЦИИ |
|
|
|
|
|
|
СТРУКТУРА ХРОМОСОМ в МИТОТИЧЕСКОМ ЦИКЛЕ: ИНТЕРФАЗНЫЙ ХРОМАТИН
ЭУХРОМАТИН |
ГЕТЕРОХРОМАТИН |
|
|
|
Факультативный |
Конститутивный |
|
(МАКСИМАЛЬНО |
(ВОЗНИКАЕТ |
(СОХРАНЯЕТСЯ на |
|
ДЕКОМПАКТИЗИ |
путем |
ПРОТЯЖЕНИИ |
|
РОВАННЫЙ, |
КОМПАКТИЗАЦИ |
ВСЕГО |
|
АКТИВНО |
И |
МИТОТИЧЕСКОГО |
|
ТРАНСКРИБИРУЕ |
ЭУХРОМАТИНА, |
ЦИКЛА): |
|
МЫЙ), |
НЕТРАНСКРИБИР |
ОКОЛОЦЕНТРОМЕРН |
|
УНИКАЛЬНЫЕ И |
УЕМЫЙ, |
ЫЕ и ТЕЛОМЕРНЫЕ |
|
МАЛОПОВТОРЯЮ |
КОМПЕНСАЦИЯ |
участки-средне- или |
|
ЩИЕСЯ |
ДОЗЫ ГЕНОВ – |
многократно |
|
ПОСЛЕДОВАТЕЛЬ |
ХРОМОСОМА Х |
||
повторяющиеся |
|||
НОСТИ. |
ГОМОГАМЕТНОГ |
||
последовательности; |
|||
|
О ПОЛА У |
||
|
предполагаемые |
||
|
ЛЮДЕЙ, фактор |
||
|
функции – |
||
|
неспецефич |
||
|
структурная , уч в |
||
|
регуляции генной |
||
|
конъюгации |
||
|
активности. |
||
|
хромосом, теломеры- |
||
|
|
||
|
|
упорядочение в |
|
|
|
объеме интерфазного |
|
|
|
ядра, защита ДНК от |
|
|
|
нуклеаз; Биотаймер. |
60. Механизмы поддержания постоянства кариотипа поколений организмов и клеток.
У организмов, размнож-ся бесполым путем, новое поколение появл в рез-те митоза, обеспеч таким образом сохранение постоянной стр-ры наследственного материала в ряду поколений клеток и орг-мов.

Рис. 3.70. Сравнение первого мейотического (редукционного) деления с митозом Профаза — спирализация хромосом, начало формирования веретена
деления; в мейозе, кроме того, происходит конъюгация гомологичных хромосом с образованием бивалентов; метафаза — в митозе в экваториальной плоскости веретена деления
располагаются отдельные хромосомы числом 2n, в мейоэе в плоскости экватора выстраивается п бивалеитов; анафаза —в митозе в результате расщепления центромер дочерние
хромосомы (бывшие сестринские хроматиды) расходятся к разным полюсам (по 2n к каждому полюсу), в мейозе разрушаются бивалеиты и гомологи расходятся к разным полюсам (по одному из каждой лары); формируется гаплоидный набор хромосом; телофаза—в митозе формируются ядра дочерних клеток, в мейозе телофа
сокращена во времени, так как не происходит полной деспирализации хромосом и клетки сразу переходят ко второму делению.
Результаты митоза — сохранение в дочерних клетках диплоидного набор хромосом (2n2с); результаты первого мейотического деления—образован клеток с гаплоидным набором двунитчатых хромосом (п2с)
При половом размножении процесс осущ с помощью гамет, вступающих в оплодотворение. При оплодотворении наследственный материал двух родительских
гамет сливается, образуя генотип организма нового поколения — зиготы. Чтобы потомки получили соответствующую программу для развития видовых и индивидуальных характеристик, они должны обладать кариотипом, которым располагало предыдущее поколение. Постоянство кариотипа в ряду поколений достигается уменьшением вдвое набора хромосом в гаметах, кот восстан до диплоидного при их оплодотворении: п +
п = 2n.
Образование гаплоидных гамет осущ в ходе гаметогенеза путем мейоза. При мейозе из клеток с диплоидным набором In образуются гаметы с гаплоидным набором хромосом п . Благодаря тому, что после однократного удвоения ДНК клетка делится дважды. В отличие от митоза в первом мейотическом делении в результате конъюгации гомологичные хромосомы объединяются в пары — биваленты. Последующее расхождение гомологов к разным полюсам веретена деления - гаплоидный набор хромосом: 2n4с → п2с. В ходе второго мейотического деления сестринские хроматиды каждой хромосомы, как и в митозе, распределяются между дочерними клетками с наследственным материалом пс. Рис. 3.71. Схема второго (эквационного) деления мейоза:
I — кл, обр в рез-те 1-го мейотического деления=двунитчатых хром (n2c);
II — кл, обр после 2-го деления мейоза и несущ гаплоидный набор однонитчатых хром
(nc)
Благодаря особенностям мейоза образуются клетки, несущ полноценный геном, в кот каждая группа сцепления представлена в единственном экземпляре (гаплоидный набор хромосом).
Сперматозоиды, проникая в яйцеклетку, вводят в нее свой ядерный наследственный материал, заключенный в гаплоидном наборе хромосом. Ядра гамет сливаются = диплоидное ядро зиготы, в кот каждая группа сцепления представлена в двойном экземпляре — отцовской и материнской хромосомами. Таким образом, мейоз и
последующее оплодотворение обеспечивают сохранение у нового поколения организмов диплоидного кариотипа, присущего всем особям данного вида.
61. Жизненный цикл клетки и его периоды.
1.Пресинтетический или постмитотический (G1) период наступает сразу же после митотического деления клетки и характеризуется активным ростом клетки и синтезом белка и РНК, благодаря чему клетка достигает нормальных размеров и восстанавливает необходимый набор органелл. G1 -период длится от нескольких часов до нескольких дней. В течение этого периода синтезируются особые "запускающие" белки, или активаторы S-периода. Они обеспечивают достижение клеткой определенного порога (точки R - рестрикции или ограничения), после которого она вступает в S-период. Контроль, осуществляемый на уровне точки R (при переходе из G1 в S), ограничивает возможность нерегулируемого размножения клеток. Проходя эту точку, клетка переключается на последующую регуляцию внутренними факторами клеточного цикла, которая обеспечивает закономерное завершение ее деления.
Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G0) для того, чтобы (в зависимости от причин остановки): -дифференцироваться и выполнять свои специфической функции -выжить в условиях недостаточности питательных веществ или факторов роста
-осуществить репарацию поврежденной ДНК. Клетки одних тканей при соответствующей стимуляции вновь способны возвращаться из периода (G0) в клеточный цикл, других - утрачивают эту способность по мере дифференцировки
2.Синтетический (S) характеризуется удвоением содержания (репликацией) ДНК и синтезом белков, в частности, гистонов, которые поступают в ядро из цитоплазмы и обеспечивают нуклеосомную упаковку вновь синтезированной ДНК. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. S- период длится у большинства клеток 8-12 часов.
3.Постсинтетический или премитотический (G2) период следует за S-периодом и продолжается вплоть до митоза (часто обозначаемого буквой М). В течение этого периода клетка осуществляет непосредственную подготовку к делению. Происходит созревание центриолей, запасается энергия, синтезируются РНК и белки (в частности, тубулин), необходимые для процесса деления. Длительность G2-периода составляет 2-4 часа. Контроль вступления клетки в митоз осуществляется двумя специальными факторами с противоположно направленными эффектами. Митоз тормозится до момента завершения репликации ДНК М-задерживающим фактором и индуцируется М-стимулирующим фактором. Действие последнего проявляется лишь в присутствии других белков - циклинов (синтезируются на протяжении всего цикла и распадаются в середине митоза).
62. Нарушения клеточного цикла. Амитоз. Эндомитоз. Политения.
Эндомитоз (от греч. endon -внутри и mitos - нить) - вариант митоза, при котором происходит удвоение числа хромосом внутри ядерной оболочки без ее разрушения и образования веретена деления. При повторных эндомитозах число хромосом в ядре может значительно увеличиваться при соответствующем кратном двум нарастании содержания в нем ДНК - полиплоидии (от греч. poly - много и ploon - складываю) и увеличении объема ядра. Полиплоидия может явиться также результатом неоконченных обычных митозов. Основной смысл развития полиплоидии заключается в усилении функциональной активности клетки.
Сходный результат достигается при образовании двуядерных клеток вследствие митотического деления, не сопровождающегося цитотомией. При последующем митотическом делении такой двуядерной клетки хромосомные наборы ядер объединяются в метафазе, приводя к образованию двух дочерних полиплоидных клеток. Наличие
полиплоидных - тетра- (4n) и октаплоидных (8n) клеток - нормальное явление в печени, эпителии мочевого пузыря, клетках концевых отделов поджелудочной и слюнных желез. Мегакариоциты (гигантские клетки костного мозга) начинают формировать кровяные пластинки лишь достигнув определенного уровня полиплоидии (16-32n) в результате нескольких эндомитозов.
Другой тип полиплоидизации-политения. При политении происходит многократная репликация нитчатых интерфазных хромосом без последующего расхождения, митоза. Клетки с политенными хромосомами никогда не делятся. В результате возникают продольные многонитчатые структуры, или политенные хромосомы.
Амитоз, или прямое деление, — это деление интерфазного ядра путем перетяжки. При амитозе веретено деления не образуется, отсутствуют все четыре фазы, свойственные митозу, спирализации хромосом не происходит, поэтому в световой микроскоп они не видны. Такое деление встречается в тканях растущего клубня картофеля, в клетках эндосперма, стенок завязи пестика, печени и хряща, роговицы глаза. Распределение клеточных органоидов, как и ДНК, осуществляется произвольно. Амитоз в отличие от митоза является самым экономичным способом деления, так как энергетические затраты при этом незначительны.
63. Бесполое размножение и его формы.
Бесполое размножение осуществляется при участии лишь одной родительской особи и происходит без образования гамет. Дочернее поколение у одних видов возникает из одной или группы клеток материнского организма, у других видов — в специализированных органах. Различают следующие способы бесполого размножения: деление, почкование, фрагментация, полиэмбриония, спорообразование, вегетативное размножение. Деление-способ бесполого размножения, характерный для одноклеточных организмов, при котором материнская особь делится на две или большее количество дочерних клеток. Можно выделить:
-простое бинарное деление (прокариоты)
-митотическое бинарное деление (простейшие, одноклеточные водоросли)- -множественное деление, или шизогонию (малярийный плазмодий, трипаносомы). Во время шизогонии сперва многократно митозом делится ядро, затем каждое из дочерних ядер окружается цитоплазмой, и формируются несколько самостоятельных организмов. Почкование-способ бесполого размножения, при котором новые особи образуются в виде выростов на теле родительской особи. Дочерние особи могут отделяться от материнской и переходить к самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом случае колонии (коралловые полипы). Фрагментация-способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды, спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации.
Полиэмбриония-способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается эмбрион (монозиготные близнецы). Вегетативное размножение-способ бесполого размножения, при котором новые особи образуются или из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.), специально предназначенных для этой формы размножения. Вегетативное размножение характерно для многих групп растений, используется в садоводстве, огородничестве, селекции растений (искусственное вегетативное размножение).
64. Митоз – цитологическая основа бесполого размножения.
Митоз (от греч. mitos - нить) является универсальным механизмом деления клеток. Митоз следует за G2-периодом и завершает клеточный цикл. Он длится 1-3 часа и обеспечивает равномерное

распределение генетического материала в дочерние клетки. Митоз включает 4 основные фазы профазу, метафазу, анафазу и телофазу.
Профаза. Эта стадия характеризуется постепенной конденсацией и спирализацией хромосом. Под микроскопом становятся различимы хромосомы в виде нитевидных структур. Видно, что каждая хромосома состоит из двух копий-дочерних хроматид, соединенных центромерой. Другим важным событием профазы является постепенное исчезновение ядрышка, содержимое которого распределяется по ядру. К полюсам клетки расходятся дочерние центриоли, и между ними формируется митотическое веретено. Митотическое веретено-это биполярная структура, которая состоит из микротрубочек.
Метафаза. На этой стадии хромосомы, прикрепленные центромерами к нитям веретена, собираются на экваторе-плоскости, расположенной посередине между полюсами веретена. Образовавшаяся метафазная пластинка представляет собой наиболее характерную особенность метафазы. Хромосомы максимально спирализованы.
Анафаза. На этой стадии каждая центромера делится пополам. Хроматиды становятся самостоятельными хромосомами, и две дочерние ценромеры устремляются к противоположным полюсам, увлекая за собой по одной из двух дочерних хромосом.
Телофаза. Два набора дочерних хромосом группируются у противоположных полюсов веретена. Происходит деспирализация хромосом. Вокруг каждого набора хромосом образуется ядерная оболочка, вновь возникает ядрышко.
Цитокинез. Разделение цитоплазмы на две обособляющиеся части. Механическим аппаратом цитокинеза является сократимое кольцо микрофиламентов, которые состоят из актина. Митотическое веретено и сократимое кольцо располагаются перпендикулярно друг другу, и веретено оказывается внутри сократимого кольца. Борозда деления разделяет плоскость митоза и образуется две генетически равноценные клетки. Цитокинез начинается в телофазе.
65. Половое размножение. Регулярные и нерегулярные формы.
Половое размножение встречается в основном у высших организмов. Оно обеспечивает значительное генетическое разнообразие и, следовательно, большую фенотипическую изменчивость потомства; организмы получают большие эволюционные возможности, возникает материал для естественного отбора.
Помимо полового размножения, существует половой процесс. Суть его в том, что обмен генетической информацией между особями происходит, но без увеличения числа особей.
Формированию гамет у многоклеточных предшествует мейоз. Половой процесс состоит в объединении наследственного материала от двух разных источников (родителей).
При половом размножении потомство генетически отличается от своих родителей, так как между родителями происходит обмен генетической информацией.
Основой полового размножения является мейоз. Родителями являются две особи – мужская и женская, они вырабатывают разные половые клетки. В этом проявляется половой диморфизм, который отражает различие задач, выполняемых при половом размножении мужским и женским организмами.
Половое размножение осуществляется через гаметы – половые клетки, имеющие гаплоидный набор хромосом и вырабатывающиеся в родительских организмах. Слияние родительских клеток приводит к образованию зиготы, из которой в дальнейшем образуется организм-потомок. Половые клетки образуются в гонадах – половых железах (в яичниках у самок и семенниках у самцов). Процесс образования половых клеток называется гаметогенезом (овогенезом у самок и сперматогенезом у самцов).
Если мужские и женские гаметы образуются в организме одной особи, то ее называют гермафродитной. Гермафродитизм бывает истинный (особь имеет гонады обоих полов) и ложный гермафродитизм (особь имеет половые железы одного типа – мужского или женского, а наружные половые органы и вторичные половые признаки обоих полов).
Партеногенез (девственное размножение) Виды партеногенеза:
1.облигатный (обязательный) партеногенез. Встречается в популяциях, состоящих исключительно из особей женского пола. При этом вероятность встречи разнополых особей минимальна 2.циклический (сезонный) партеногенез (у тлей, дафний, коловраток). Встречается в популяциях, которые исторически вымирали в больших количествах в определенное время года. У этих видов партеногенез сочетается с половым размножением. При этом в летнее время существуют только самки, которые откладывают два вида яиц – крупные и мелкие. Из крупных яиц партеногенетически появляются самки, а из мелких – самцы, которые оплодотворяют яйца, лежащие зимой на дне. Из них появляются исключительно самки; 3.факультативный (необязательный) партеногенез. Встречается у общественных насекомых (ос,
пчел, муравьев). В популяции пчел из оплодотворенных яиц выходят самки (рабочие пчелы и царицы), из неоплодотворенных – самцы (трутни).
Гиногенез (у костистых рыб и некоторых земноводных). Сперматозоид проникает в яйцеклетку и лишь стимулирует ее развитие. Ядро сперматозоида при этом с ядром яйцеклетки не сливается и погибает, а источником наследственного материала для развития потомка служит ДНК ядра яйцеклетки.
Андрогенез. В развитии зародыша участвует мужское ядро, привнесенное в яйцеклетку, а ядро яйцеклетки при этом гибнет. Яйцеклетка дает лишь питательные вещества своей цитоплазмы. Полиэмбриония. Зигота (эмбрион) делится на несколько частей бесполым способом, каждая из которых развивается в самостоятельный организм. Встречается у насекомых (наездников), броненосцев. У броненосцев клеточный материал первоначально одного зародыша на стадии бластулы равномерно разделяется между 4–8 зародышами, каждый из которых в дальнейшем дает полноценную особь.
К этой категории явлений можно отнести появление однояйцовых близнецов у человека. У одноклеточных организмов выделяют две формы полового размножения – копуляцию и конъюгацию.
При конъюгации (например, у инфузорий) специальные половые клетки не образуются. При этом процессе не происходит увеличения количества особей, поэтому говорят о половом процессе, а не о половом размножении. Однако происходит обмен (рекомбинация) наследственной информацией, поэтому потомки генетически отличаются от своих родителей.
При копуляции (у простейших) происходят образование половых элементов и их попарное слияние. При этом две особи приобретают половые различия и полностью сливаются, образуя зиготу. Происходят объединение и рекомбинация наследственного материала, поэтому особи генетически отличны от родительских.
66. Цитологические основы полового размножения. Мейоз, как специфический процесс при формировании половых клеток.
Мейоз состоит из двух последовательных клеточных делений, которые соответственно называются мейоз I и мейоз II. В первом делении происходит уменьшение числа хромосом в два раза, поэтому первое мейотическое деление называют редукционным. Во втором делении число хромосом не изменяется; такое деление называют эквационным.
Интерфаза Предмейотическая интерфаза отличается от обычной интерфазы тем, что процесс
репликации ДНК не доходит до конца. Таким образом, деление клетки начинается на синтетической стадии клеточного цикла. Поэтому мейоз образно называют преждевременным митозом. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с.
Первое деление мейоза (редукционное деление, или мейоз I)
Сущность редукционного деления заключается в уменьшении числа хромосом в два раза: из исходной диплоидной клетки образуется две гаплоидные клетки с двухроматидными хромосомами (в состав каждой хромосомы входит 2 хроматиды).
Профаза 1 состоит из ряда стадий:
Лептотена. Хромосомы видны в световой микроскоп в виде клубка тонких нитей. Раннюю лептотену, когда нити хромосом видны еще очень плохо,
называют пролептотена.
Зиготена. Происходит конъюгация гомологичных хромосом. Гомологичные хромосомы – это хромосомы, сходные между собой в морфологическом и генетическом отношении. При конъюгации образуются биваленты. Каждый бивалент – это относительно устойчивый комплекс из одной пары гомологичных хромосом. Гомологи удерживаются друг около друга с помощью белковых синаптонемальных комплексов. Количество бивалентов равно гаплоидному числу хромосом. Иначе биваленты называются тетрады, так как в состав каждого бивалента входит 4 хроматиды.
Пахитена. Хромосомы спирализуются, хорошо видна их продольная неоднородность. Завершается репликация ДНК. Завершается кроссинговер.
Диплотена. Гомологичные хромосомы в бивалентах отталкиваются друг от друга. Они соединены в отдельных точках, которые называются хиазмы.
Диакинез (стадия расхождения бивалентов). Отдельные биваленты располагаются на периферии ядра.
Метафаза I. Ядерная оболочка разрушается (фрагментируется). Формируется веретено деления. Далее происходит метакинез – биваленты перемещаются в экваториальную плоскость клетки.
Анафаза I. Гомологичные хромосомы, входящие в состав каждого бивалента, разъединяются, и каждая хромосома движется в сторону ближайшего полюса клетки. Разъединения хромосом на хроматиды не происходит. Процесс распределения хромосом по дочерним клеткам называется сегрегация хромосом.
Телофаза I. Гомологичные двухроматидные хромосомы полностью расходятся к полюсам клетки. В норме каждая дочерняя клетка получает одну гомологичную хромосому из каждой пары гомологов. Формируются два гаплоидных ядра, которые содержат в два раза меньше хромосом, чем ядро исходной диплоидной клетки. Каждое гаплоидное ядро содержит только один хромосомный набор, то есть каждая хромосома представлена только одним гомологом. Содержание ДНК в дочерних клетках составляет 2с.
Вбольшинстве случаев (но не всегда) телофаза I сопровождается цитокинезом. Интеркинез-это короткий промежуток между двумя мейотическими делениями. Отличается от интерфазы тем, что не происходит репликации ДНК, удвоения хромосом и удвоения центриолей.
Второе деление мейоза (эквационное деление, или мейоз II)
Входе второго деления мейоза уменьшения числа хромосом не происходит.
Сущность эквационного деления заключается в образовании четырех гаплоидных клеток с однохроматидными хромосомами (в состав каждой хромосомы входит одна хроматида).