

BIO_-_vsyo
.pdfпосттранскрипц-х изменения РНК, в рез-те к-рых из пре-иРНК образ зрел иРНК. Во время процессинга происх кэпирование-присоед-е к 5' концу мол-лы метилированного гуанозинтрифосфата, к-ый назыв полиаденилиров-е-расщепл-е растущего транскрипта в определ-м месте и добавл-е к 3' концу в точке разреза последоват-ти поли-А, состоящей из 100-300 остатков аденозина; сплайсинг-вырез-е интронов (неинформативных уч-в) и сшив- е экзонов (информативныз уч-в)
17. Регуляция экспрессии генов у прокариот. Индукция синтеза катаболических ферментов(Lac-оперон).
Регул экспрессии генов у прокариот наиболее распространена регул на уровне транскрипции (очень экономна). Гипотеза оперона. Оперон-совок-ть тесно сцепл-х структ- х генов прокариот вместе с уч-ком-оператором, регулирующим их транскрипцию. К оперону также относят регуляторн уч-ки:промотор и терминатор, к-ые участв в управл транскрипцией.Промотор-уч-к ДНК,обозначающ оконч-е транскрипции.РНК-полимераза, дойдя до терминатора, заканч транскрипцию и покид ДНК. Оператор-уч-к ДНК, с к-рым может специфически связыв-ся белок-репрессор.Инф-цию о белке-репрессоре содержит ген-регулятор, к-рый не вход в состав оперона.Связавшись с оператором, репрессор останавл транскрипцию, т.к. препятств передвиж-ю РНК-полимеразы вдоль ДНК. Lac-оперон включ в себя 3 структ-х гена, к-рые несут инф-цию о ферментах , расщепл-х лактозу.В отс-е лактозы ферм-ты, её расщепляющие, не синтезир-ся. Их синтез останавл на стадии транскрипции.Транскрипцию блокир белок-репрессор,к-рый связ с оператором. Индуктором, к-рый включ синтез ферм-в, явл лактоза Когда в кл-ке появл лактоза, она связыв с белком-репрессором и перевод его в неакт форму. Неакт репрессор теряет спость связыв-ся с ДНК и уход с оператора. РНК-полимераза получ возм-ть транскрибировать структ гены. В рез-те транскрипции образ иРНК, содержащая три структ-х гена. На кажд гене происход трансляция, в рез-те к-рой синтезир-ся ферм-ты, расщепляющие лактозу.По мере расщепл-я лактозы её конц падает, белок-репрессор освобожд-ся от лактозы,переход в акт форму,приобрет спос-ть связ с ДНК,садится на оперон и останавл трансляцию.
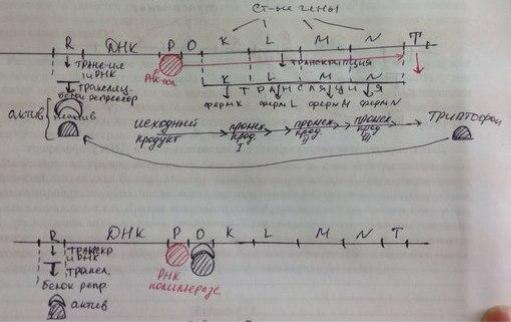
18. Регуляция экспрессии генов у прокариот. Репрессия синтеза анаболических ферментов(trp-оперон).
Регул экспрессии генов у прокариот наиболее распространена регул на уровне транскрипции(оч экономна). Гипотеза оперона. Оперон-совок-ть тесно сцепл-х структ-х генов прокариот вместе с уч-ком-оператором, регулирующим их транскрипцию. К оперону также относят регуляторн уч-ки:промотор и терминатор, к-ые участв в управл транскрипцией.Промотор-уч-к ДНК,обозначающ оконч-е транскрипции.РНК-полимераза, дойдя до терминатора, заканч транскрипцию и покид ДНК. Оператор-уч-к ДНК, с к-рым может специфически связыв-ся белок-репрессор.Инф-цию о белке-репрессоре содержит ген-регулятор, к-рый не вход в состав оперона.Связавшись с оператором, репрессор останавл транскрипцию, т.к. препятств передвиж-ю РНК-полимеразы вдоль ДНК. trp-оперон содерж 4 структ гена, на к-рых запис инф-ция о ферментах, катализир-х синтез триптофана. Белок-репрессор синтезир-ся в неакт форме и не может связыв-ся с ДНК,поэтому возможна транскрипция структ-х генов и дальнейш синтез ферментов, вырабатыв-х триптофан. По мере синтеза триптофана его конц в кл-ке возраст. Избыт триптофан соед с белком-репрессором и перевод его в акт форму. Репрессор приобрет спос-ть связ ДНК, садится на оператор и останавл транскрипцию, а след-но и синтез триптофана. В процессе жизнед-ти кл-ки триптофан расход-ся, белок-репрессор освобожд от триптофана, переход в неакт сост-е, теряет спос-ть связ с ДНК и покид оператор. Транскрипуия и последующ процессы синтеза возобновл-ся.
19. Общие принципы генетического контроля экспрессии генов.
Важнейшим фактором регуляции генной акт-ти явл элементы генома, отвечающие за синтез регуляторных белков,— гены-регуляторы. Соединяясь с определ-ными нуклеотидными последовательностями ДНК, предшествующими структурной части регулируемого гена,—операторами, белки-регуляторы способств или препятств соедин-ю РНК-полимеразы с промотором. Если белок-регулятор взаимод с оператором, занимающим часть промотора или располож-м между ним и структурной частью гена, то это не дает возможности РНК-полимеразе соедин-ся с промоторной последоват-тью и осуществить транскрипцию. Такой белок называют репрессором, и в этом случае осуществл-ся негативный контроль экспрессии гена со стороны гена-регулятора. Если
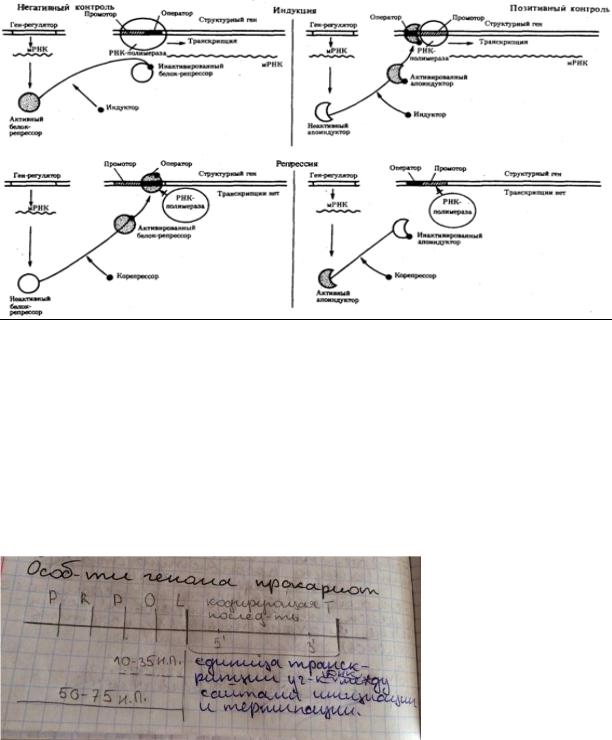
промотор облад слабой спос-тью соединяться с РНК-полимеразой, а ему предшествует область, узнаваемая белком-регулятором, присоедин-е последнего непоср-но перед промотором к молекуле ДНК облегч связыв-е РНК-полимеразы с промотором, вслед за чем следует транскрипция. Такие белки назыв активаторами (или апоиндукторами), а контроль экспрессии гена со стороны гена-регулятора — позитивным
20. Роль регуляторных белков в регуляции генной активности (репрессоры, активаторы).
Все виды РНК-полимераз узнают свои промоторы при помощи регулят-х белков, называемых факторами транскрипции. Факторы транскрипции связыв-ся с ДНК в обл промотора. (один из них – ТАТА-фактор, к-рый связыв-ся с последоват-тью ТАТААТ в области промотора, что способств присоед-ю РНК-полимеразы II). Активаторы вызыв транскрипцию, репрессоры подавл её.
21. Организация генома прокариот.

По химической организации материала наследсвенности и изменчивости экариоты и прокариоты принципиально не отличаются друг от друга. Генетический материал у них представлен ДНК. Общим для них является и принцип записи генетической информации, а также генетический код. Одни и те же аминокилосты шифруются у про- и эукариот одинаковыми кодонами. Принципиально одинаковым образом у названных типов клеток осуществляется и использование наследсвенной информации, хранящейся в ДНК. Сначала она транскрибируется в нуклеотидную последовательность молекулы мРНК, а затем транслируется в ак последовательность пептида на рибосомах с участием тРНК. Особенности: -наследственный материал прокариота содержится главным образом в единственной кольцевой молекуле ДНК. Она распологается непосредсвенно в цитопламе клетки, где также находятся необходимые для экспрессии генов тРНК и ферменты, часть из которых заключена в рибосомах. –гены прокариот непрерывны –в процессе транскрипции участвует только один фермент – рнк-полимераза . –у прокариот 3 вида регуляторных участков ДНК:1)промоторный, для связи с РНК-полимеразой, 10-35 н.п левее сайта инициации; 2)терминаторный, отвечает за завершение транскрипции и высвобождения транскрипта; 3)операторный, сцеплен с промотором или перекрывается с ним.
22. Организация генома эукариот. По химической организации материала наследсвенности и изменчивости экариоты и прокариоты принципиально не отличаются друг от друга. Генетический материал у них представлен ДНК. Общим для них является и принцип записи генетической информации, а также генетический код. Одни и те же аминокилосты шифруются у про- и эукариот одинаковыми кодонами. Принципиально одинаковым образом у названных типов клеток осуществляется и использование наследсвенной информации, хранящейся в ДНК. Сначала она транскрибируется в нуклеотидную последовательность молекулы мРНК, а затем транслируется в ак последовательность пептида на рибосомах с участием тРНК. Особенности: -гены преривистые( информативные участки-экзоны и неинформативные участки-интроны);- экспрессию генов обеспечивает три вида рнк-полимераза 1,2,3.-регуляторные участки обычно расположены левее сайта инициации. Два вида:-промоторный( для связи с рнкполимеразой 100 н.п левее сайта инициации могут быть за сотни и даже тысячи н.п левее сайта инициации);-энхансеры(от 100 до 20 000 н.п левее стимулир. Транскр-цию при связ). –прицип построения генома эукариот три уровня:-генный, хромосомный, геномный.
– регуляция экспрессии генов происходит на всех этапах экспрессии генов. Механизмы регуляции многообразны и невероятно сложны.

23. Неклеточные формы жизни. Вирусы.
Этот термин применяют для обозначения уникальных представителей живой природы, не имеющих клеточного (эукариотического или прокариотического) строения и обладающих облигатным внутриклеточным паразитизмом, т.е. которые не могут жить без клетки.
Вирусы принципиально отличаются от всех других организмов. Назовем их важнейшие особенности.
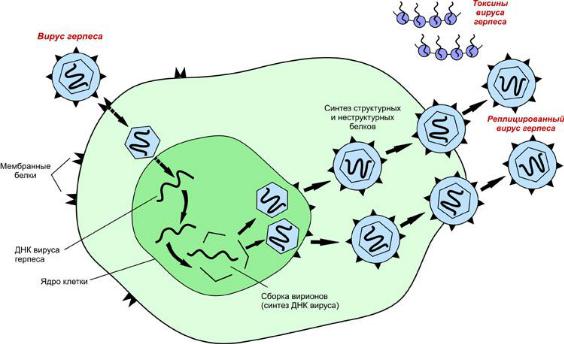
1.Тело не имеет клеточного строения.2. Они могут существовать только как внутриклеточные паразиты и не могут размножаться вне клеток тех организмов, в которых они паразитируют.3. Содержат лишь один тип нуклеиновых кислот - либо РНК, либо ДНК (все клеточные организмы содержат и ДНК, и РНК одновременно). Нет рибосом.4. Нет обмена веществ. Используется энергия, получаемая за счет обмена веществ в клетках хозяина. Имеют очень ограниченное число ферментов, используют обмен веществ хозяина, его ферменты, энергию, полученную при обмене веществ в клетках хозяина.5. Зрелые вироспоры ("споры" вирусов) могут существовать вне клетки хозяина, в этот период они не обнаруживают никаких признаков жизни.Долгие годы продолжался спор: вирусы - это живые существа или часть неживой природы. Невозможность существования и размножения вирусов вне клетки, их способность к самосборке и кристаллизации говорили о том, что вирус ведет себя как "неживая" материя. После установления природы гена и обнаружения в вирусах генетического материала, присущего живым организмам, вирусы стали относить к живой природе. Генетический аппарат вирусов представлен различными формами нуклеиновых кислот, такого разнообразия нет ни у одной из других форм жизни. У всех живых организмов, кроме вирусов, генетический аппарат состоит из двунитевой молекулы дезоксирибонуклеиновой кислоты (ДНК), а рибонуклеиновая кислота (РНК), выполняющая в клетках роль переносчика информации, всегда однонитевая. У вирусов же существуют все возможные варианты устройства генетического аппарата: одно- и двунитевая РНК, одно- и двунитевая ДНК. При этом и вирусная РНК, и вирусная ДНК могут быть либо линейными, либо замкнутыми в кольцо. Основные этапы взаимодействия вируса с клеткой хозяина.
1.Адсорбция - пусковой механизм, связанный со взаимодействием специфических рецепторов вируса и хозяина.
2.Проникновение - путем слияния суперкапсида с мембраной клетки или путем эндоцитоза
(пиноцитоза).
3.Освобождение нуклеиновых кислот - “раздевание” нуклеокапсида и активация нуклеиновой кислоты.
4.Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и их работа на воспроизводство вируса.
5.Сборка вирионовассоциация реплицированных копий вирусной нуклеиновой кислоты с капсидным белком.
6.Выход вирусных частиц из клетки, приобретения суперкапсида оболочечными вирусами.
24. Ген – функциональная единица наследственности. Эволюция представлений о гене. История изучения генов.
1909г-Иогансен впервые ввел термин ген. Вейсман предположил, что гены находятся в хромосомах. 1911-Морган доказал, что гены расположены в хромосомах. Хромосомная теория наследственности. 1953-Уотсон и Крик-двойна спираль ДНК (объясняет свойства днк) 50егг-бум развития молекулярной биолгии(после решения проблемы генетического кода). Центральный постулат молекулярной биологии: в живых системах поток информации возможен только в одном направлении днк-рнк-белок. В 1961 году-обратная транскрипция. 1970 года-экспериментальное док-ва наличия фермента-обратная транскриптаза. Далее, обнаружили прерывистость гена. Оказалось, что у эукариот геныэто чередование смысловых(экзоны) и бессмысленных(интронов) участков ДНК. Первый продукт траанскрипции-гетерогенная РНК имет экзонно-интронную структуру(прерывистую). Сплайсинг-вырезание из первичного транскрипта интронов и сшивание экзонов. Вторичный экзонный продукт транскрипции-зрелая иРНК. Альтернативный сплайсинг-из одного первичного РНК-транскрипта в разных тканях образуется несколько разных по длинне зрелых –Рнк. Т.о., одна и та же ДНКпоследовательность может кодировать несколько белковых продуктов. Вывод: на современном этапе развития биологии трудно дать исчерпывающие определение гена.
Ген-экспрессирующая единица генома, включающая единицу транскрипции и регуляторные участки. (ген прокариот непрерывеню) Возможность проявления гена в виде признака зависит от других генов гомологичной хромосомы и от условий внешней среды.У всех организмов одного вида каждый конкретный ген расположен в одном и том же месте - локусе - строго определенной хромосомы.
25. История изучения структуры гена.
1909г-Иогансен впервые ввел термин ген. Вейсман предположил, что гены находятся в хромосомах. 1911-Морган доказал, что гены расположены в хромосомах. Хромосомная теория наследственности. 1953-Уотсон и Крик-двойна спираль ДНК (объясняет свойства

днк) 50егг-бум развития молекулярной биолгии(после решения проблемы генетического кода). Центральный постулат молекулярной биологии: в живых системах поток информации возможен только в одном направлении днк-рнк-белок. В 1961 году-обратная транскрипция. 1970 года-экспериментальное док-ва наличия фермента-обратная транскриптаза. Далее, обнаружили прерывистость гена. Оказалось, что у эукариот геныэто чередование смысловых(экзоны) и бессмысленных(интронов) участков ДНК. Первый продукт траанскрипции-гетерогенная РНК имет экзонно-интронную структуру(прерывистую). Сплайсинг-вырезание из первичного транскрипта интронов и сшивание экзонов. Вторичный экзонный продукт транскрипции-зрелая иРНК. Альтернативный сплайсинг-из одного первичного РНК-транскрипта в разных тканях образуется несколько разных по длинне зрелых –Рнк. Т.о., одна и та же ДНКпоследовательность может кодировать несколько белковых продуктов. Вывод: на современном этапе развития биологии трудно дать исчерпывающие определение гена.
26. Сравнительная характеристика геномов прокариот и эукариот.
По химической организации материала наследсвенности и изменчивости экариоты и прокариоты принципиально не отличаются друг от друга. Генетический материал у них представлен ДНК. Общим для них является и принцип записи генетической информации, а также генетический код. Одни и те же аминокилосты шифруются у про- и эукариот одинаковыми кодонами. Принципиально одинаковым образом у названных типов клеток осуществляется и использование наследсвенной информации, хранящейся в ДНК. Сначала она транскрибируется в нуклеотидную последовательность молекулы мРНК, а затем транслируется в ак последовательность пептида на рибосомах с участием тРНК. Особенности: -наследственный материал прокариота содержится главным образом в единственной кольцевой молекуле ДНК. Она распологается непосредсвенно в цитопламе клетки, где также находятся необходимые для экспрессии генов тРНК и ферменты, часть из которых заключена в рибосомах. –гены прокариот непрерывны –в процессе транскрипции участвует только один фермент – рнк-полимераза . –у прокариот 3 вида регуляторных участков ДНК:1) промоторный, для связи с РНК-полимеразой, 10-35 н.п левее сайта инициации; 2) терминаторный, отвечает за завершение транскрипции и высвобождения транскрипта; 3) операторный, сцеплен с промотором или перекрывается с ним.

27.Международная программа «Геном человека».
28.Основные этапы программы «Геном человека». Значимость проекта для современной медицины.
Важнейшие геномные проекты XX и XXI веков.Проект по расшифровке генома— международный научно-исследовательский проект, главной целью которого было определить последовательность нуклеотидов, которые составляют ДНК и идентифицировать 20—25 тыс. генов в человеческом геноме.Проект начался в 1990 году, под руководством Джеймса Уотсона под эгидой Национальной организации здравоохранения США. В 2000 году был выпущен рабочий черновик структуры генома, полный геном — в 2003 году, однако и сегодня дополнительный анализ некоторых участков ещё не закончен. Частной компанией «Celera Genomics » был запущен аналогичный параллельный проект, завершённый несколько ранее международного. Основной объём секвенирования был выполнен в университетах и исследовательских центрах США, Канады и Великобритании. Кроме очевидной фундаментальной значимости, определение структуры человеческих генов является важным шагом для разработки новых медикаментов и развития других аспектов здравоохранения.Хотя целью проекта по расшифровке генома человека является понимание строения генома человеческого вида, проект также фокусировался и на нескольких других организмах, среди которых бактерии, насекомые, такие как мушка дрозофила, и млекопитающие, например, мышь.Изначально планировалось определение последовательности более трёх миллиардов нуклеотидов, содержащихся в гаплоидном человеческом геноме. Затем несколько групп объявили о попытке расширить задачу до секвенирования диплоидного генома человека.Геном любого отдельно взятого организма (исключая однояйцевых близнецов и клонированных животных) уникален, поэтому определение последовательности человеческого генома в принципе должно включать в себя и секвенирование многочисленных вариаций каждого гена. Однако, в задачи проекта «Геном человека» не входило определение последовательности всей ДНК, находящейся в человеческих клетках; а некоторые гетерохроматиновые области (в общей сложности около 8 %) остаются несеквенированными до сих пор. Последовательность человеческой ДНК сохраняется в базах данных, доступных любому пользователю через Интернет. Национальный центр биотехнологической информации США (и его партнёрские организации в Европе и Японии) хранят геномные последовательности в базе данных известной как GenBank, вместе с последовательностями известных и гипотетических
генов и белков.Процесс идентификации границ генов и других мотивов в необработанных последовательностях ДНК называется аннотацией генома (англ.) и относится к области биоинформатики. Эту работу при помощи компьютеров выполняют люди, но они делают её медленно и, чтобы удовлетворять требованиями высокой пропускной способности проектов секвенирования геномов, здесь также всё шире используют специальные компьютерные программы.Другая, часто упускаемая из виду цель проекта «Геном человека» — исследование этических, правовых и социальных последствий расшифровки генома. Важно исследовать эти вопросы и найти наиболее подходящие решения до того, как они станут почвой для разногласий и политических проблем.Все люди имеют в той или иной степени уникальные геномные последовательности. Поэтому данные, опубликованные проектом «Геном человека», не содержат точной последовательности геномов каждого отдельного человека. Это комбинированный геном небольшого количества анонимных доноров. Почти все цели, которые ставил перед собой проект, были достигнуты быстрее, чем предполагалось. Проект по расшифровке генома человека был закончен на два года раньше, чем планировалось. Проект поставил разумную, достижимую цель секвенирования 95 % ДНК. Исследователи не только достигли её, но и превзошли собственные предсказания, и смогли секвенировать 99,99 % человеческой ДНК. Проект не только превзошёл все цели и выработанные ранее стандарты, но и продолжает улучшать уже достигнутые результаты.
29. Организация генома человека.
Ядро-95% ДНК. Общая протяженность ДНК ядра 1,5-2 метра разделена на 23 фрагмента. В митохондриях-5% ДНК. Около 1000 МТХ содержится в одной клетке. МТХ ДНК не содержит интронов. Физический размер генома человека 3*10^9 нуклеотидных пар. Только 3-5% ДНК кодируют белки, 97% ДНХ “отдыхает”(издержка эволюции). Двадцать две пары аутосом, две половые хромосомы Х и Y, а также митохондриальная ДНК человека содержат вместе примерно 3,1 млрд пар оснований. Известно, что Y-хромосома содержит 78 генов. Из них 60 контролирует сперматогенез, а остальные определяют другие функции(сравните с 2 тыс генов в Х-хромосоме). В человеческом геноме найдено множество различных последовательностей, отвечающих за регуляцию гена. Под регуляцией понимается контроль экспрессии гена (процесс построения матричной РНК по участку молекулы ДНК). Обычно это короткие последовательности, находящиеся либо рядом с геном, либо внутри гена. Иногда они находятся на значительном расстоянии от гена (энхансеры)
30. Понятие о геномике и новый взгляд на эволюцию.
Геномика — раздел молекулярной генетики, посвящённый изучению генома и генов живых организмов. Есть несколько разделов геномики. Структурная геномика — содержание и организация геномной информации. Имеет целью изучение генов с известной структурой для понимания их функции, а также определение пространственного строения максимального числа «ключевых» белковых молекул. Функциональная геномика — реализация информации, записанной в геноме, от гена — к признаку. Сравнительная геномика (эволюционная) — сравнительные исследования содержания и организации геномов разных организмов. Сравнение генов привело к парадоксальным выводам: эволюция от низших форм к высшим сопряжена с разбавлением генома-на единицу длинны ДНК приходится все меньше информации о структуре белков и все больше информации ни о чем. Это большая загадка биологической эволюции. Ф. Крик лишнюю ДНК назвал эгоистической, считал издержкой эволюции. Примером является ALU-повторы. Локализация этого повтора в ДНК человека от 500 тыс.
до 1 млн сайтов. Переход к человеку-взрывообразное нарастание ALU-повтором. Ученые связывают элементы познания с ALU-повторами и считают их личными генами человека. Сравнительная геномика демонстрирует поразительное сходство геномов различных организмов ( Геном человека высоко консервативен). Вывод: процесс эволюции сопровождается структурными реорганизациями генома. Важную роль играли мутации, но не меньшее значение имели и имеют перестановки блоков геном(фрагментов генома)
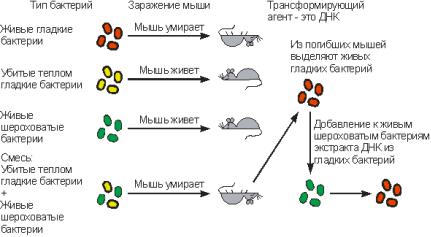
31. Экспериментальные доказательства генетической роли нуклеиновых кислот. Трансформация.
Трансформация – направленный перенос и встраивание в генетический аппарат клетки небольшого фрагмента чужеродной ДНК. Она происходит без участия вирусов – бактериофагов. Посредством генетической рекомбинации часть трансформирующей молекулы ДНК может обмениваться с частью хромосомной ДНК донора. Известно, что бактерия Pneumococcus pneumonie имеет несколько форм. Вирулентность ее определяется наличием мукополисахаридной капсулы на поверхности клетки, которая защищает бактерию от воздействия со стороны организма – хозяина. Капсула – слой полипептидов или полисахаридов, липидов или гетерополисахаридов и до 90% воды, расположенных поверх клеточной стенки и выполняющий функции осмотического барьера, защиты от высыхания и механических повреждений. В результате размножившиеся бактерии убивают зараженное животное. Бактерии этого штамма (S-штамм) образуют гладкие колонии. Авирулентные формы не имеют защитной капсулы и образуют шероховатые колонии (R-штамм). Микробиолог Ф. Гриффитс в 1928 году инъецировал мышам культуру живого пневмококка R-штамма вместе с S-штаммом, убитым высокой температурой равной 65 градусов Цельсия. (см. рисунок).
Спустя некоторое время ему удалось выделить из зараженных мышей живые пневмококки, обладающие капсулой. Таким образом, оказалось, что свойство убитого пневмококка – способность образовывать капсулы – перешло к живой бактерии, то есть, произошла трансформация этих клеток. От этого превращения клеток и возник сам этот термин. Поскольку признак наличия капсулы является наследственным, то следовало предположить, что какая-то часть наследственного вещества от бактерий штамма – S перешла к клеткам штамма – R. Таким образом, стало ясно, что от одного штамма бактерий к другому возможна передача наследственного начала, однако химическая природа его не была обнаружена. Химическая природа вещества, трансформирующего