

Плазматическая мембрана. Барьерно-рецепторная и транспортная системы клетки
Плазматическая мембрана, или плазмолемма (membrana cellularis), среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой, а следовательно, и со всеми веществами и стимулами, воздействующими на клетку.
Химический состав плазматической мембраны. Основу плазмолеммы составляет липопротеиновый комплекс. Она имеет толщину около 10 нм и, таким образом, является самой толстой из клеточных мембран.
Снаружи от плазмолеммы располагается надмембранный слой - глико-каликс (glycocalyx). Толщина этого слоя около 3-4 нм, он обнаружен практически у всех животных клеток, но степень его выраженности различна. Гликокаликс представляет собой ассоциированный с плазмолеммой гли-копротеиновый комплекс, в состав которого входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы (см. рис. 4.2). При использовании специальных методов выявления полисахаридов (краситель рутениевый красный) показано, что они образуют структуру, подобную «чехлу» поверх плазматической мембраны.
В гликокаликсе могут располагаться белки, не связанные с билипидным слоем. Как правило, это белки-ферменты, участвующие во внеклеточном расщеплении различных веществ, таких как углеводы, белки, жиры и др.
Функции плазматической мембраны - это разграничение цитоплазмы с внешней средой, рецепции и транспорта различных веществ как внутрь клетки, так и из нее.
Рецепторные функции связаны с локализацией на плазмолемме специальных структур, участвующих в специфическом «узнавании» химических и физических факторов. Клеточная поверхность обладает большим набором компонентов - рецепторов, определяющих возможность специфических реакций с различными агентами. Рецепторами на поверхности клетки могут служить гликопротеиды и гликолипиды мембран (см. рис. 4.2). Считается, что такие чувствительные к отдельным веществам участки могут быть разбросаны по всей поверхности клетки или собраны в небольшие зоны. Существуют рецепторы биологически активных веществ - гормонов, медиаторов, специфических антигенов разных клеток или белков и др.
С плазмолеммой связана локализация специфических рецепторов, отвечающих за такие важные процессы, как взаимное распознавание клеток и развитие иммунитета. Так, в составе плазмолеммы всех клеток находится молекула гистосовместимости I класса (гликопротеин), которая состоит из: а) интегрального трансмембранного белка, часть которого находится в цитоплазме, другая - пронизывает плазмолемму, и последняя, наиболее длинная часть молекулы, находится в гликокаликсе; б) периферического мембранного белка с малой молекулярной массой; в) короткой молекулы белка, которая нековалентно связывается с петлями внеклеточной части интегрального трансмембранного белка. Именно последняя часть молекулы (пептид из 9 аминокислот) является фрагментом нормального белка клетки данной особи. Он и распознается как «свой» клетками иммунной системы человека. В случае мутации на месте белка гистосовместимости появляется белок с иной структурой молекулы (например, кодируемый вирусом), и в ответ на это возникает иммунная реакция со стороны организма, направленная на уничтожение данной клетки. Этот механизм сохраняет генетическую индивидуальность клеток, следовательно, и организма.
В плазмолемме светочувствительных клеток животных расположена специальная система фоторецепторных белков (родопсин), с помощью которых световой сигнал превращается в химический, что, в свою очередь, приводит к генерации электрического импульса.
Выполняя транспортную функцию, плазмолемма обеспечивает пассивный перенос ряда веществ, например воды, ряда ионов и некоторых низкомолекулярных соединений. Другие вещества проникают через мембрану путем активного переноса против градиента концентрации с затратой энергии за счет расщепления АТФ. Так транспортируются многие органические молекулы (сахара, аминокислоты и др.). Эти процессы могут быть сопряжены с транспортом ионов, в них участвуют белки-переносчики.
Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и даже их агрегаты, а часто и крупные частицы попадают внутрь клетки в результате процесса эндо-цитоза (рис. 4.3). Эндоцитозформально разделяют на фагоцитоз (захват и
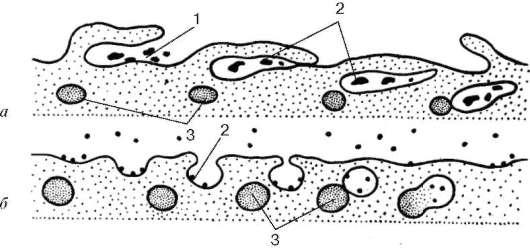
Рис. 4.3. Эндоцитоз. Разные типы образования пиноцитозных пузырьков (а, б):
1 - сорбция частиц на поверхности плазматической мембраны; 2 - погружение
частиц в цитоплазму; 3 - первичные лизосомы
поглощение клеткой крупных частиц, например, бактерий или фрагментов других клеток) и пиноцитоз (захват отдельных молекул и макромолекуляр-ных соединений).
Пиноцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней.
В дальнейшем эндоцитозные пузырьки, или эндосомы, могут сливаться друг с другом, расти, и в их внутренней полости, кроме поглощенных веществ, начинают обнаруживаться гидролитические ферменты (гидролазы), поступающие сюда излизосом (см. ниже). Эти ферменты расщепляют биополимеры до мономеров, которые в результате активного транспорта через мембрану пузырька переходят в гиалоплазму. Таким образом, поглощенные молекулы внутри мембранных вакуолей, образовавшихся из элементов плазмолеммы, подвергаются внутриклеточному пищеварению.
При фагоцитозе клетка, например макрофаг, после того, как к ее плазмо-лемме прикрепляется бактерия, формирует длинные цитоплазматические отростки, которые охватывают бактерию, и макрофаг постепенно поглощает бактерию с образованием фагосомы.
Плазмолемма принимает участие в выведении веществ из клетки (экзоци-тоз). В этом случае внутриклеточные продукты (белки, мукополисахариды, липопротеиды и др.), заключенные в вакуоли или пузырьки и отграниченные от гиалоплазмы мембраной, подходят к плазмолемме. В местах контактов плазмолемма и мембрана вакуоли сливаются, и содержимое вакуоли поступает в окружающую среду.
Процесс эндоцитоза и экзоцитоза осуществляется при участии связанной с плазмолеммой системы фибриллярных компонентов цитоплазмы, таких как микротрубочки и сократимые микрофиламенты. Последние, соединяясь с определенными участками плазмолеммы, могут, изменяя свою длину, втягивать мембрану внутрь клетки, что приводит к отделению от плазмолеммы эндоцитозных вакуолей. Часто, непосредственно примыкая к плазмолемме, микрофиламенты образуют сплошной кортикальный слой.
Плазмолемма многих клеток животных может образовывать выросты. У ряда клеток такие выросты включают в свой состав специальные компоненты цитоплазмы (микротрубочки, фибриллы), что приводит к развитию органелл движения -ресничек, жгутиков и др.
Наиболее часто встречаются на поверхности многих животных клеток микроворсинки. Это выросты цитоплазмы, ограниченные плазмолеммой, имеющие форму цилиндра с закругленной вершиной. Микроворсинки характерны для клеток эпителия, но обнаруживаются и у клеток других тканей. Диаметр микроворсинок около 100 нм. Число и длина их различны у разных типов клеток. Возрастание числа микроворсинок приводит к резкому увеличению площади клеточной поверхности. Это особенно важно для клеток, участвующих во всасывании. Так, в кишечном эпителии на