

1 Мм2 поверхности насчитывается до 2?108 микроворсинок.
Межклеточные соединения
Плазмолемма принимает активное участие в образовании специальных структур - межклеточных контактов, илисоединений (junc-tiones intercellulares), обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур (рис. 4.4).
Общим для этих клеток является то, что на их поверхности располагаются специальные углеводные части интегральных белков, глико-протеидов, которые специфически взаимодействуют и соединяются с соответствующими белками на поверхности соседних клеток.
Межклеточные соединения делятся на простые и сложные.
Простое межклеточное соединение ( junctio intercellularis simplex) - сближение плазмолемм соседних клеток на расстояние 15-20 нм (рис. 4.5). При этом происходит взаимодействие слоев гликокалик-са соседних клеток. С помощью
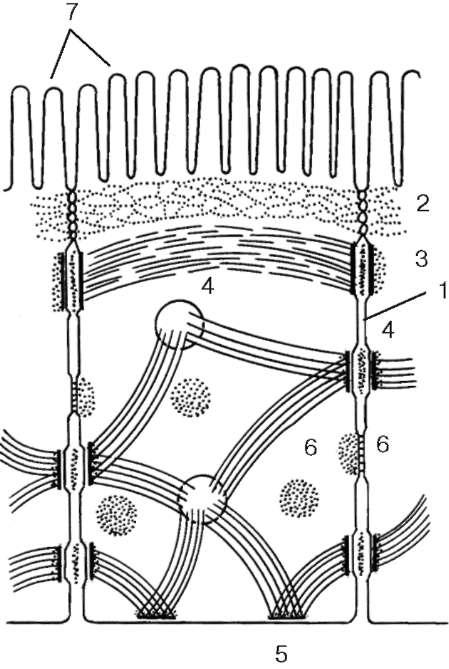
Рис. 4.4. Расположение различных межклеточных соединений в клетках кишечного эпителия (схема):
1 - простое соединение; 2 - плотное соединение (замыкающая зона); 3 - адгезивный поясок (поясок слипания); 4 - десмосома (пятно сцепления); 5 - полудес-мосома; 6 - щелевое (коммуникационное) соединение; 7 - микроворсинки
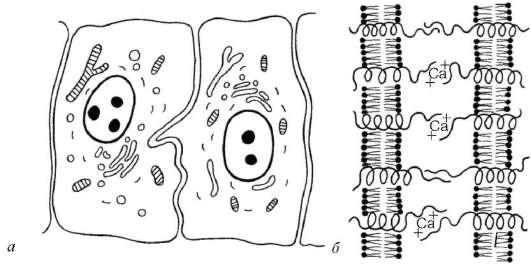
Рис. 4.5. Простое межклеточное соединение (схема):
а - простое соединение двух эпителиальных клеток; б - связывание интегральными гликопротеидами (интегринами и кадгеринами) плазматических мембран соседних клеток
белков-рецепторов, входящих в состав гликокаликса, происходит распознавание клеток, возникших из общего зачатка, и объединение их в пласты. Например, Е-кадгерины участвуют в образовании контактов только между эпителиальными клетками, обеспечивая их соединение практически по всей поверхности контактирующих клеток.
Сложные межклеточные соединения представляют собой небольшие парные специализированные участки плазматических мембран двух соседних клеток. Они подразделяются на замыкающие (изолирующие), сцепляющие (заякоривающие) и коммуникационные (объединяющие) соединения.
К замыкающим относится плотное соединение (замыкающая зона - zonu-la occuludens). В этом соединении принимают участие специальные интегральные белки, расположенные на поверхности соседних клеток, образующие подобие ячеистой сети (рис. 4.6).
Эта ячеистая сеть окружает в виде пояска весь периметр клетки, соединяясь с такой же сетью на поверхности соседних клеток. Данная область непроницаема для макромолекул и ионов, и, следовательно, она замыкает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды. Этот тип соединений характерен для клеток однослойных эпителиев и эндотелия некоторых сосудов.
К сцепляющим, или заякоривающим, соединениям относятся адгезивный поясок (поясок слипания) и десмосома. Общим для этой группы соединений является то, что к участкам плазматических мембран со стороны цитоплазмы подходят фибриллярные элементы цитоскелета (актиновые филаменты,

Рис. 4.6. Плотное соединение (замыкающая зона):
а - расположение плотного соединения (вставочная пластинка) на клетках кишечного эпителия; б - трехмерная схема участка плотного соединения. 1 - микроворсинки
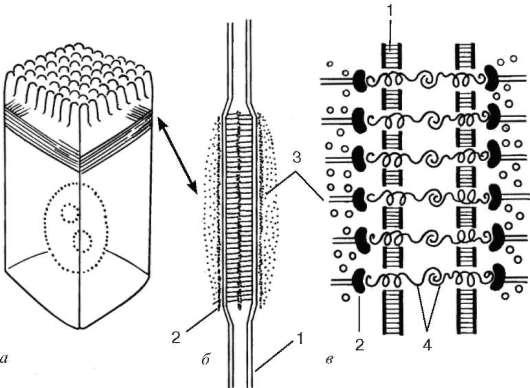
Рис. 4.7. Адгезивный поясок (поясок слипания):
а - расположение его в клетке; б - вид на срезе; в - схема молекулярной организации. 1 - плазмолемма; 2 - слой белков сцепления; 3 - актиновые микрофиламенты; 4 - связующие гликопротеиды
промежуточные филаменты и спектрин) и связываются с мембранами в области соединения соседних клеток.
Адгезивный поясок, или поясок слипания (zonula adherens), - парное образование в виде лент, каждая из которых опоясывает апикальные части соседних клеток и обеспечивает в этой области их прилипание друг к другу (рис. 4.7). Здесь клетки связаны друг с другом интегральными гликопротеи-дами, к которым со стороны цитоплазмы и той и другой клетки примыкает слой примембранных белков, включающих характерный белок винкулин. К этому слою подходит и связывается с ним пучок актиновых микрофила-ментов. Взаимодействие актиновых микрофиламентов с актинсвязывающи-ми белками во многих соседствующих клетках может привести к изменению рельефа всего эпителиального пласта.
К сцепляющим соединениям может быть отнесен фокальный контакт, характерный для фибробластов. В этом случае клетка соединяется не с соседней клеткой, а с элементами внеклеточного субстрата. В образовании фокального контакта также принимают участие актиновые микрофила-менты. К сцепляющим межклеточным соединениям относятся идесмосомы (рис. 4.8).
Десмосома, или пятно слипания (macula adherens). Это парные структуры, представляющие собой небольшую площадку или пятно диаметром около 0,5 мкм. Со стороны цитоплазмы к плазматической мембране прилежит слой белков, в состав которого входят десмоплакины. В этот слой со стороны цитоплазмы внедряются пучки промежуточных филаментов. С внешней стороны плазмолеммы соседних клеток в области десмосом соединяются
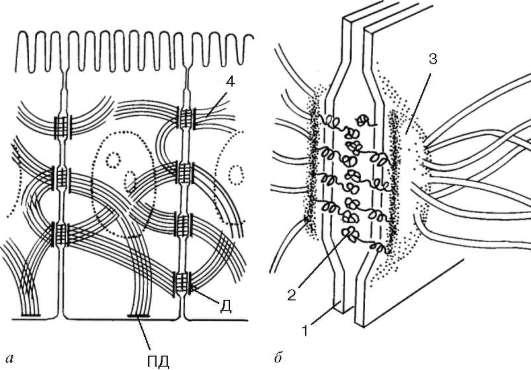
Рис. 4.8. Десмосома:
а - расположение в клетке; б - схема ультраструктуры. 1 - плазмолемма; 2 - десмо-глеиновый слой; 3 - слой десмоплакина; 4 - промежуточные филаменты. Д - десмосома; ПД - полудесмосома
с помощью трансмембранных белков - десмоглеинов. Например, каждая клетка эпидермиса кожи может иметь до нескольких сотен десмосом.
Функциональная роль десмосом заключается главным образом в механической связи между клетками. Десмосомы связывают друг с другом клетки в различных эпителиях, в сердечной и гладких мышцах. Полудесмосомы связывают эпителиальные клетки с базальной мембраной.
Коммуникационные соединения в клетках животных представлены щелевыми соединениями и синапсами (рис. 4.9).
Щелевое соединение, или нексус (nexus), представляет собой область протяженностью 0,5-3 мкм, где плазмолеммы разделены промежутком в 2-3 нм (см. рис. 4.9). Со стороны цитоплазмы никаких специальных примембран-ных структур в данной области не обнаруживается, но в структуре плазмо-лемм соседних клеток друг против друга располагаются специальные белковые комплексы (коннексоны), которые образуют каналы из одной клетки в другую. Этот тип соединения встречается во всех группах тканей.
Функциональная роль щелевого соединения заключается в переносе ионов и мелких молекул (молекулярная масса 2?103) от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексусы.
Синаптические соединения, или синапсы (synapsis). Этот тип соединений характерен для нервной ткани и встречается в специализированных участках контакта как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора (например, нервно-мышечные, нервно-эпителиальные синапсы).
Синапсы - участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому (см. главу 10).

Рис. 4.9. Щелевое (коммуникационное) соединение:
1 - коннексон; 2 - плазмолемма
Вакуолярная система
Эндоплазматическая сеть
Эндоплазматическая сеть (эндо-плазматический ретикулум) была открыта К. Р. Портером в 1945 г. Этот компонент вакуолярной системы клетки представляет собой совокупность вакуолей, плоских мембранных мешков или трубчатых образований, создающих трехмерную мембранную сеть. В состав сети входят гранулярные и агранулярные участки, которые могут чередоваться.
Гранулярная эндоплазматическая сеть (reticulum endoplasmicum granulosum) на ультратонких срезах представлена замкнутыми мембранами, которые на сечениях образуют уплощенные мешки, цистерны, трубочки.
Диаметр цистерн значительно варьирует и в зависимости от функциональной активности клетки колеблется от 20 нм до несколько микрометров. Отличительной чертой мембран гранулярной эндоплазматической сети является то, что они со стороны гиалоплазмы покрыты многочисленными рибосомами (рис. 4.10).
Гранулярная эндоплазматическая сеть имеет разное строение. Для малоспециализированных клеток или для клеток с низкой метаболической активностью характерно наличие редких и разрозненных цистерн. Если возникают локальные скопления гранулярной эндоплазматической сети, то это свидетельствует об активном синтезе секреторных белков. Так, в клетках печени и некоторых нервных клетках гранулярная эндоплазматическая сеть собрана в отдельные зоны. В клетках поджелудочной железы гранулярная эндоплазмати-ческая сеть в виде плотно упакованных друг около друга мембранных цистерн занимает базальную и околоядерную зоны клетки. Рибосомы, связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков - ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Белки, накапливающиеся в полостях эндоплазматической сети, могут, минуя гиалоплазму, транспортироваться в вакуоли комплекса Гольджи, где они модифицируются и входят в состав либо лизосом, либо секреторных гранул, содержимое которых остается изолированным от гиалоплазмы мембраной. Внутри канальцев или вакуолей гранулярной эндоплазматической
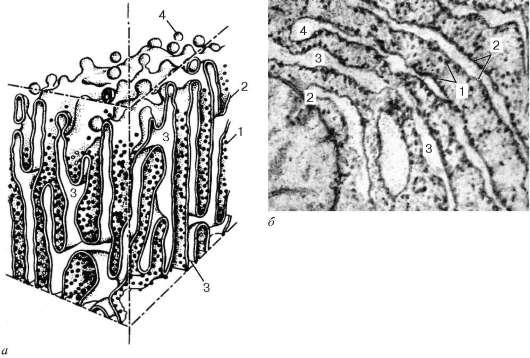
Рис. 4.10. Строение гранулярной эндоплазматической сети: а - схема; б - электронная микрофотография участка среза эпителиальной клетки печени. 1 - рибосомы; 2 - пластинки; 3 - внутренние полости цистерн; 4 - отщепляющиеся мембранные пузырьки, лишенные рибосом
сети происходит модификация белков, например, связывание их с сахарами (первичное глюкозилирование).
В гранулярной эндоплазматической сети на ее рибосомах происходит синтез мембранных интегральных белков, которые встраиваются в толщу мембраны. Здесь же со стороны гиалоплазмы идет синтез липидов и их встраивание в мембрану. В результате этих двух процессов наращиваются сами мембраны эндоплазматической сети и другие компоненты вакуоляр-ной системы клетки.
Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран.
Агранулярная (гладкая) эндоплазматическая сеть (reticulum endoplasmaticum nongranulosum) также представлена мембранами, образующими мелкие вакуоли, трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50-100 нм. Гладкая эндоплаз-матическая сеть возникает и развивается на основе гранулярной эндоплаз-матической сети. В отдельных участках гранулярной эндоплазматической сети образуются новые липопротеидные мембранные участки, лишенные рибосом. Эти участки могут разрастаться, отщепляться от гранулярных мембран и функционировать как самостоятельная вакуолярная система.
Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих стероиды, например, в эндокринных клетках коркового вещества надпочечников, в эпителиальных клетках извитых семенных канальцев.
Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
В поперечнополосатых мышечных волокнах гладкая эндоплазматиче-ская сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани (см. главу 9).
Очень важна роль гладкой эндоплазматической сети в дезактивации различных вредных для организма веществ за счет их окисления с помощью ряда специальных ферментов. Особенно четко она проявляется в клетках печени. Так, при некоторых отравлениях в клетках печени появляются ацидофильные зоны (не содержащие РНК), сплошь заполненные гладким эндо-плазматическим ретикулумом.