

Ядерная оболочка
Ядерная оболочка (tegmentum nucleare), или кариолемма, состоит из внешней ядерной мембраны (m. nuclearis externa) ивнутренней мембраны оболочки (m. nucle-aris interna), разделенных перинуклеарным пространством (рис. 4.26). Ядерная оболочка содержит многочисленные ядерные поры (pori nucleares).
Из многих свойств и функциональных нагрузок ядерной оболочки следует подчеркнуть ее роль как барьера, отделяющего содержимое ядра от цитоплазмы, ограничивающего свободный доступ в ядро крупных агрегатов биополимеров, регулирующего транспорт макромолекул между ядром и цитоплазмой.
Мембраны оболочки ядра в морфологическом отношении не отличаются от остальных внутриклеточных мембран. В общем виде оболочка ядра может быть представлена как полый двухслойный мешок, отделяющий содержимое ядра от цитоплазмы.
Наружная мембрана оболочки ядра, непосредственно контактирующая с цитоплазмой клетки, имеет ряд структурных особенностей, позволяющих отнести ее к собственно мембранной системе эндоплазматической сети: на ней со стороны гиалоплазмы расположены многочисленные полирибосомы, а сама наружная мембрана может прямо переходить в мембраны эндоплаз-матической сети. Одной из важных функций оболочки ядра следует считать ее участие в создании внутриядерного порядка - в фиксации хромосомного материала в трехмерном пространстве ядра. В интерфазе часть хроматина структурно связана с внутренней мембраной оболочки ядра. Эта связь опосредуется с помощью фиброзной ядерной пластинки (ламины), с которой связываются фибриллы хроматина.
Наиболее характерными структурами оболочки ядра являются ядерные поры. Они образуются за счет слияния наружной и внутренней мембран обо-
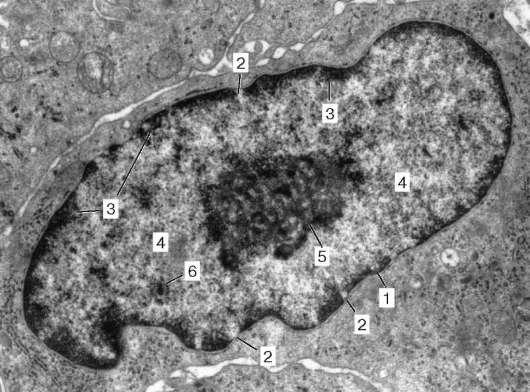
Рис. 4.26. Строение ядра интерфазной клетки:
1 - оболочка ядра (наружная и внутренняя мембраны, перинуклеарное пространство); 2 - комплекс ядерной поры; 3 - гетерохроматин; 4 - эухроматин; 5 - ядрышко; 6 - межхроматиновые гранулы РНК. Электронная микрофотография, увеличение 12 000
лочки ядра. Формирующиеся при этом округлые сквозные отверстия поры имеют диаметр около 90 нм. Эти отверстия в ядерной оболочке заполнены сложно организованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют комплексом ядерной поры (complexus pori nuclearis). Последний имеет октагональную симметрию. По границе отверстия в наружной и внутренней мембранах оболочки ядра располагаются по 8 белковых субъединиц, которые составляют белковые кольца ядерной поры (наружное и внутреннее). От наружного кольца поры в сторону цитоплазмы отходят длинные филаменты. От внутреннего кольца поры в глубь ядра также отходят филаменты, образуя структуру, подобную корзинке.
Комплекс ядерной поры в функциональном отношении представляет собою сложную систему, которая активно участвует не только в рецепции транспортируемых макромолекул (белков и нуклеопротеидов), но и собственно в актах их переноса (транслокации), при которых используется АТФ. В состав каждого комплекса ядерной поры входит несколько сотен различных белков.
Число ядерных пор зависит от метаболической активности клеток: чем интенсивнее синтетические процессы в клетках, тем больше пор в оболочке ядра. Так, у эритробластов (клеток-предшественников ядерных эритроцитов) низших позвоночных животных во время интенсивного синтеза и накопления гемоглобина в оболочке ядра обнаруживаются около 30 пор на
1 мкм2 поверхности. После того как эти процессы заканчиваются, в ядрах зрелых клеток - эритроцитов - прекращается синтез ДНК и РНК и количество пор в оболочке ядра снижается до 5 на 1 мкм2 поверхности. В оболочке ядра зрелых сперматозоидов поры не обнаруживаются. В среднем в оболочке ядра соматической клетки обнаруживается несколько тысяч поро-вых комплексов.
4.3. ВОСПРОИЗВЕДЕНИЕ КЛЕТОК 4.3.1. Клеточный цикл и его регуляция
Делению клеток предшествует редупликация хромосом вследствие синтеза ДНК. Это правило является общим для про- и эукариотических клеток. Время существования клетки от одного деления до другого называется клеточным циклом (cyclus cellularis).
Во взрослом организме высших позвоночных клетки различных тканей и органов имеют неодинаковую способность к делению. Встречаются популяции клеток, полностью потерявшие свойство делиться. Это большей частью специализированные, дифференцированные клетки (например, зернистые лейкоциты крови). В организме есть постоянно обновляющиеся ткани - различные эпителии, кроветворные ткани. В таких тканях существует часть клеток, которые постоянно делятся, заменяя стареющие или погибающие клетки (например, клетки базального слоя покровного эпителия, клетки крипт кишечника, кроветворные клетки костного мозга). Многие клетки, не размножающиеся в обычных условиях, приобретают вновь это свойство при процессах репаратив-ной регенерации органов и тканей. В гистогенезе большинство клеток после определенного числа делений переходит в гетеросинтетическую интерфазу, которая включает время роста, дифференцировки, функционирования, старения и смерти. В целом это характеризует жизненный цикл клетки.
При изучении клеточного цикла встречаются как диплоидные (2 с), так и тетраплоидные (4 с) и интерфазные клетки с промежуточным количеством ДНК. Это объясняется особенностями цикла размножения клеток. Весь клеточный цикл состоит из четырех отрезков времени: собственно митоза (М), пресинтети-ческого (G1), синтетического (S) и постсинтетического (G2) периодов интерфазы (рис. 4.27).
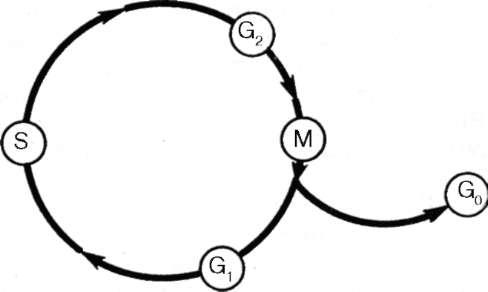
Рис. 4.27. Клеточный цикл (схема). Пояснения в тексте
В G1-периоде, наступающем сразу после деления, клетка имеет диплоидное содержание ДНК в ядре (2 с). После деления в периоде G1 в дочер-
них клетках общее содержание белков и РНК вдвое меньше, чем в исходной родительской клетке. В периоде G1наблюдается рост клеток главным образом за счет накопления клеточных белков, что обусловлено увеличением количества РНК в клетке, и подготовка клетки к синтезу ДНК.
Обнаружено, что подавление синтеза белка или иРНК в G1-периоде предотвращает наступление S-периода, так как в течение G1-периода происходят синтезы ферментов, необходимых для образования предшественников ДНК (например, нуклеотидфосфокиназ), ферментов метаболизма РНК и белка. При этом резко повышается активность ферментов, участвующих в энергетическом обмене.
В следующем, S-периоде, происходит удвоение количества ДНК в ядре и соответственно удваивается число хромосом. В ядрах разных клеток, находящихся в S-периоде, можно обнаружить разные количества ДНК - от 2 до 4 с, что отражает постепенное накопление ДНК по мере прохождения клеткой синтетического периода клеточного цикла. S-период является узловым в клеточном цикле. Без синтеза ДНК неизвестно ни одного случая вступления клеток в митотическое деление.
Единственным исключением является второе деление созревания половых клеток в мейозе, когда между двумя делениями нет синтеза ДНК.
В S-периоде уровень синтеза РНК возрастает соответственно увеличению количества ДНК, достигая своего максимума в С2-периоде.
Постсинтетический (G2) период называется также премитотическим. В данном периоде синтезируется иРНК, необходимая для митоза. Среди синтезирующихся в это время белков особое место занимают тубулины - белки митотического веретена.
В конце G2-периода или в митозе по мере конденсации митотических хромосом синтез РНК резко снижается и полностью прекращается во время митоза. Синтез белка во время митоза понижается до 25 % исходного уровня и затем в последующих периодах достигает своего максимума в G2-периоде, в общем повторяя характер синтеза РНК.
В растущих тканях растений и животных всегда есть клетки, которые находятся вне цикла. Такие клетки принято называть клетками G0-периода. Это клетки, которые после митоза не вступают в пресинтетический период (G1). Именно они представляют собой так называемые покоящиеся, временно или окончательно переставшие размножаться клетки. В некоторых тканях такие клетки могут находиться длительное время, не изменяя особенно своих морфологических свойств: они сохраняют способность к делению. Это, например, камбиальные клетки (стволовые в кроветворной ткани). Чаще потеря (хотя бы и временная) способности делиться сопровождается специализацией и дифференцировкой. Такие дифференцирующиеся клетки выходят из цикла, но в особых условиях могут снова входить в цикл. Например, большинство клеток печени находится в G0-периоде; они не синтезируют ДНК и не делятся. Однако при удалении части печени у экспериментальных животных многие клетки начинают подготовку к митозу, переходят к синтезу ДНК и могут митотически делиться. В других случаях, например в эпидермисе кожи, после выхода клетки из цикла
размножения они дифференцируются, выполняют свои защитные функции, а затем погибают (ороговевшие клетки покровного эпителия). Многие клетки теряют полностью способность возвращаться в митотический цикл. Так, например, нейроны головного мозга и кардио-миоциты постоянно находятся в G0-периоде клеточного цикла (до смерти организма).
Регуляция вхождения и выхода клеток из клеточного цикла находится под контролем особой системы белковых факторов. Было обнаружено множество факторов роста (ФР), стимулирующих клетки к размножению, к пролиферации. Так, например, ФР из тромбоцитов стимулирует размножение клеток соединительной ткани, гормон эритро-поэтин вызывает размножение пред-
шественников эритроцитов, гормон прогестерон стимулирует пролиферацию клеток молочной железы и т. п.
Разные ФР передают сигналы на синтез специальных внутриклеточных белков, образующих каскад протеинкиназ (фосфорилаз), связанных с запуском клеточного цикла.
В состав этих белков, факторов, стимулирующих митоз, входит комплекс, состоящий из двух субъединиц: регуляторной (белок циклин) и каталитической (циклин-зависимая протеиназа).
У млекопитающих в реализации всего клеточного цикла участвуют 9 различных циклинов и 7 циклин-зависимых киназ (ЦЗК). При этом для перехода от одного периода клеточного цикла к другому используются разные циклины (D, E, A, B и др.) и разные ЦЗК (рис. 4.28). Так, например, переход от интерфазного ядра в G2-периоде непосредственно к митозу определяется фактором, состоящим из циклинов А/В и протеин-зависимой киназы 1.
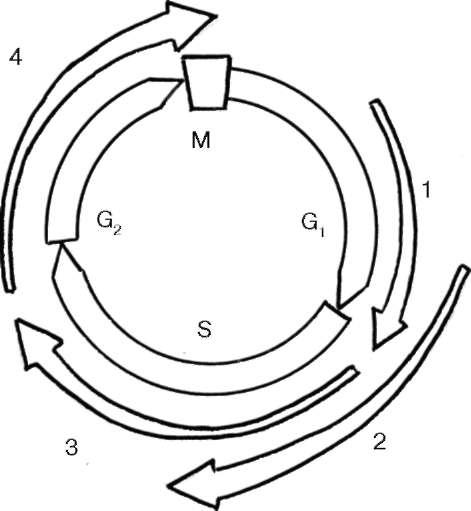
Рис. 4.28. Участие различных циклинов и циклин-зависимых киназ в клеточном цикле млекопитающих: 1 - циклин D+ЦЗК 4, ЦЗК 6; 2 - циклин Е+ЦЗК 2; 3 - циклин А+ЦЗК 2; 4 - циклин В/А+ЦЗК 1
Деление клеток: митоз
Митоз (mitosis), кариокинез, или непрямое деление, - универсальный способ деления любых эукариотических клеток. При этом редуплицирован-ные и конденсированные хромосомы переходят в компактную форму мито-тических хромосом, образуется веретено деления, участвующее в сегрегации и переносе хромосом (ахроматиновый митотический аппарат), происходят расхождение хромосом к противоположным полюсам клетки и деление тела клетки (цитокинез, цитотомия). Процесс непрямого деления клеток приня-
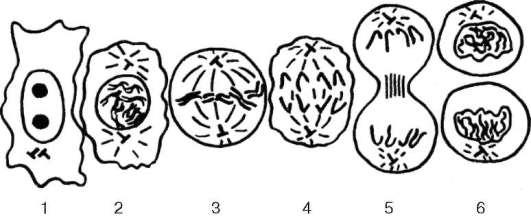
Рис. 4.29. Митоз клетки (схема):
1 - интерфаза; 2 - профаза; 3 - метафаза; 4 - анафаза; 5 - телофаза; 6 - ранняя интерфаза
то подразделять на несколько основных фаз: профаза, метафаза, анафаза, телофаза (рис. 4.29).
Профаза. После окончания S-периода количество ДНК в интерфазном ядре равно 4 с, так как произошло удвоение хромосомного материала. Однако морфологически регистрировать удвоение числа хромосом в этом периоде не всегда удается. Это связано с тем, что в профазе сестринские хромосомы тесно соприкасаются и взаимно спирализуются одна относительно другой. Тем не менее в профазе каждая из хромосом двойная, что является результатом их редупликации в S-периоде клеточного цикла. Позднее хромосомы в каждой такой паре начинают обособляться, раскручиваться. Сестринские хромосомы в митозе отчетливо выявляются в конце профазы, когда видно, что общее их число в начинающей делиться клетке равно 4 n. Следовательно, уже в начале профазы хромосомы состояли из двух сестринских хромосом, или хроматид. Число их (4 n) в профазе точно соответствует количеству ДНК (4 с).
Параллельно конденсации хромосом в профазе происходят исчезновение и дезинтеграция ядрышек в результате инактивации рибосомных генов в зоне ядрышковых организаторов.
Одновременно с этим в середине профазы начинается разрушение ядерной оболочки: исчезают ядерные поры, оболочка распадается сначала на фрагменты, а затем на мелкие мембранные пузырьки.
В это время меняются и структуры, связанные с синтезом белка. Происходит уменьшение количества гранулярной эндоплазматической сети, она распадается на короткие цистерны и вакуоли, число рибосом на ее мембранах резко падает. Значительно (до 25 %) редуцируется число полисом как на мембранах, так и в гиалоплазме, что является признаком общего снижения уровня синтеза белка в делящихся клетках.
Второе важнейшее событие при митозе тоже происходит во время профазы - это образование веретена деления. В профазе репродуцировавшиеся в S-периоде центриоли начинают расходиться к противоположным полюсам клетки. К каждому полюсу отходит по двойной центриоли, или диплосоме. По мере расхождения диплосом начинают формироваться микротрубоч-
ки, отходящие от периферических участков одной из центриолей каждой диплосомы.
Сформированный в метафазе аппарат деления в животных клетках имеет веретеновидную форму и состоит из нескольких зон: двух зон центросфер с центриолями внутри них и промежуточной между ними зоны волокон веретена. Во всех этих зонах имеется большое число микротрубочек (рис. 4.30).
Микротрубочки в центральной части этого аппарата, в собственном веретене деления, так же как микротрубочки центросфер, возникают в результате полимеризации тубулинов в зоне центриолей. Эти микротрубочки достигают кине-тохоров, расположенных в области центромерных перетяжек хромосом, и связываются с ними. В веретене деления различают два типа микротрубочек: идущие от полюса к центру веретена и хромосомные, соединяющие хромосомы с одним из полюсов.
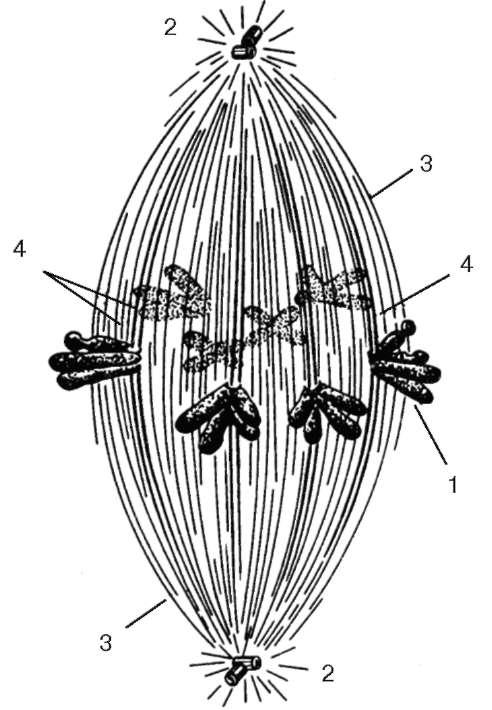
Рис. 4.30. Строение митотического веретена (схема):
1 - хромосомы; 2 - клеточный центр; 3 - центриолярные микротрубочки; 4 - кинетохорные микротрубочки
Метафаза занимает около трети времени всего митоза. Во время метафа-зы заканчивается образование веретена деления, а хромосомы выстраиваются в экваториальной плоскости веретена, образуя так называемую экваториальную (метафазную) пластинку хромосом, или материнскую звезду. К концу метафазы завершается процесс обособления друг от друга сестринских хроматид. Их плечи лежат параллельно друг другу, между ними хорошо видна разделяющая их щель. Последним местом, где контакт между хрома-тидами сохраняется, является центромер (первичная перетяжка).
Анафаза. Хромосомы все одновременно теряют связь друг с другом в области центромер и синхронно начинают удаляться друг от друга по направлению к противоположным полюсам клетки. Скорость движения хромосом равномерная, она может достигать 0,2-0,5 мкм/мин. Анафаза - самая короткая стадия митоза (несколько процентов от всего времени), но за это время происходит ряд событий. Главными из них являются обособление двух идентичных наборов хромосом и перемещение их в противоположные концы клетки. Расхождение хромосом по направлению к полюсам происходит одновременно с расхождением самих полюсов.
Показано, что расхождение хромосом связано с укорочением (деполимеризацией) микротрубочек в районе кинетохоров хромосом и с работой
белков-транслокаторов, перемещающих хромосомы. Дополнительное расхождение полюсов в анафазе обеспечивается за счет скольжения относительно друг друга межполюсных микротрубочек, которое обеспечивается работой другой группы белков-транслокаторов.
Телофаза начинается с остановки разошедшихся диплоидных (2 n) наборов хромосом (ранняя телофаза) и завершается, когда возникает реконструкция нового интерфазного ядра (поздняя телофаза, ранний G1-период) и происходит разделение исходной клетки на две дочерние (цитокинез, цитотомия). В ранней телофазе хромосомы, не меняя своей ориентации (центромерные участки - к полюсу, теломерные - к центру веретена), начинают деконденсироваться и увеличиваться в объеме. В местах их контактов с мембранными пузырьками цитоплазмы образуется новая ядерная оболочка. После замыкания ядерной оболочки начинается формирование новых ядрышек. Клетка переходит в новый G1-период клеточного цикла.
Важное событие телофазы - разделение клеточного тела - цитото-мия, или цитокинез, который происходит путем образования перетяжки в результате впячивания плазматической мембраны внутрь клетки. При этом в подмембранном слое цитоплазмы располагаются сократимые элементы типа актиновых миофиламентов, ориентированные циркулярно в зоне экватора клетки. Сокращение филаментов приводит к впячиванию плазматической мембраны в области этого кольца, что завершается разделением клетки на две.
Аномалии клеточного деления
При повреждении митотического аппарата (действие холода или агентов, вызывающих деполимеризацию тубулинов) может произойти или задержка митоза в метафазе, или рассеивание хромосом. При нарушениях репродукции центриолей могут возникать многополюсные и асимметричные митозы и т. п. Нарушения цитотомии приводят к появлению клеток с гигантскими ядрами или многоядерных полиплоидных клеток. Это связано с подавлением образования актиновых микрофиламентов, участвующих в формировании клеточной перетяжки в конце телофазы.
Полиплоидия - образование клеток с повышенным содержанием ДНК. Такие полиплоидные клетки появляются в результате полного отсутствия или незавершенности отдельных этапов митоза. Появление полиплоидных соматических клеток может наблюдаться в норме при блокаде деления клеточного тела. В печени взрослых млекопитающих встречаются, кроме диплоидных, тетра- и октаплоидные (4 n и 8 n) клетки, а также двуядерные клетки разной степени плоидности.
Процесс полиплоидизации этих клеток происходит следующим образом. После S-периода клетки, обладающие 4 с количеством ДНК, вступают в митотическое деление, проходят все его стадии, включая телофазу, но не приступают к цитотомии. Таким образом, образуется двуядерная клетка (2x2 n). Если она снова проходит S-период, то оба ядра в такой клетке будут содержать по 4 с ДНК и 4 n хромосом. Такая двуядерная клетка входит в митоз, на стадии метафазы происходит объединение хромосомных
наборов (общее число хромосом равно 8 n), а затем - нормальное деление, в результате которого образуются две тетраплоидные клетки. Этот процесс попеременного появления двуядерных и одноядерных клеток приводит к появлению ядер с 8 n, 16 n и даже 32 n количеством хромосом. Подобным способом образуются полиплоидные клетки в печени, в эпителии мочевого пузыря, в пигментном эпителии сетчатки, в ацинарных отделах слюнных и поджелудочной желез, мегакариоциты костного мозга.
Необходимо отметить, что полиплоидизация соматических клеток характерна для специализированных, дифференцированных клеток и не встречается при генеративных процессах, таких как эмбриогенез (исключая провизорные органы) и образование половых клеток; нет полиплоидии среди стволовых клеток.
Процесс митотического деления клеток очень чувствителен к действию самых разнообразных факторов. Наиболее часто встречается остановка митоза на стадии метафазы. Это происходит в результате изменений веретена деления. Многие вещества, останавливающие митоз, например, такие цитостатики, как колхицин и колцемид, препятствуют полимеризации тубулинов. В результате этого новые микротрубочки веретена не образуются, а готовые полностью разбираются. При этом митотические хромосомы собираются в центре клетки, но не образуют метафазную пластинку, а располагаются без всякого порядка (К-митоз). К сходным результатам приводит действие на клетку ингибиторов синтеза АТФ (динитрофенол, олигомицин) и ряда ядовитых веществ (мер-каптоэтанол). Если действие этих факторов кратковременное, то возможны восстановление микротрубочек веретена и клеточное деление. При умеренных воздействиях клетки могут не погибнуть, а без митоза вступать в следующий клеточный цикл. В этом случае неразошедшие-ся хромосомы деконденсируются, образуются новая ядерная оболочка и новое, но уже тетраплоидное ядро, которое переходит в G1-фазу. Так возникают полиплоидные клетки при действии колхицина.
К аномалиям деления клеток относятся и многополюсные митозы. В этом случае в метафазе образуется не биполярное веретено, а веретено с тремя или четырьмя полюсами. Такая аномалия связана с нарушениями функций центриолей: диплосома распадается на две активные моноцентриоли. Эти изменения могут происходить спонтанно (что характерно для опухолевых клеток) или после воздействия различных ингибиторов митотического деления. Такие аномальные трех- и четы-рехполюсные митотические фигуры могут вступать в анафазу и участвовать в расхождении хромосом к полюсам, вслед за чем может наступить цитотомия с образованием 3 или 4 клеток. В этих случаях не происходит равномерного распределения хромосом, а образовавшиеся клетки содержат случайные и уменьшенные наборы хромосом. Клетки с ненормальным числом хромосом называют анэуплоидными. Эти клетки обычно быстро погибают.
Нарушения митотического деления могут быть связаны со структурными изменениями самих хромосом. Так, воздействие различными формами лучистой энергии (ультрафиолетовый свет, рентгеновские лучи и т. п.) или разными алкилирующими соединениями (иприт, цитоста-тики) может привести к нарушениям структуры хромосом и изменениям хода митоза. В результате таких воздействий возникают так называемые хромосомные аберрации. Это могут быть делеции - потери участков хромосом, инверсии - перестановка участков хромосом, транслокации - перенос участков с одной хромосомы на другие.
При разрыве хромосомы та ее часть, которая не несет центромеры, не участвует в хромосомном делении, отстает от основной массы хромосом и случайно оказывается в одной из дочерних клеток. Такой фрагмент хромосомы в интерфазе покрывается собственной ядерной оболочкой (возникает дополнительное микроядро). Ясно, что при этом обе дочерние клетки будут анэуплоидными.
В других случаях в результате объединения двух поврежденных хромосом возникает одна хромосома, но с двумя центромерами, которые растягиваются к противоположным полюсам. При этом между двумя группами хромосом в анафазе и в телофазе виден «мост», возникает растянутая аберрантная хромосома.