

Глава 12 взаємини організмів із середовищем
План
12.1 Навколишнє середовище
12.2 Умови середовища
12.3 Організм і середовище
12.4 Популяція й навколишнє середовище
12.5 Екосистеми
Кожен організм перебуває в різноманітних взаємних зв'язках з факторами навколишнього середовища, як абіотичними (геофізичними, геохімічними), так і біотичними (живими організмами того ж й інших видів). Ці надорганізмені зв'язки на різних рівнях інтеграції вивчає екологія – наука про «господарство» природи. Термін «екологія» в 1866 р. запропонував видатний німецький біолог Э.Геккель. Досліджувані рівні – це взаємини особин (аутекологія), популяцій (популяційна екологія або демекологія) і співтовариств організмів (синекологія, системна екологія) з їхнім оточенням.
12.1 Навколишнє середовище
До навколишнього середовища відносяться ті елементи середовища, з якими даний організм вступає в прямі або непрямі відносини. За характером цих відносин навколишнє середовище можна розділити на різні «шари».
A. Сприймане, інформативне середовище. Це специфічний для даного виду «фільтрат» навколишнього світу. Людина не бачить ультрафіолетове світло, а бджола червоний. Ультразвукові локаційні крики кажанів не сприймаються людиною, а нічні метелики прекрасно їх чують.
Б. Мінімальне середовище охоплює безпосередньо необхідні для життя енергетичні й матеріальні ресурси навколишнього світу (світло, їжа, життєвий простір і т.д.) незалежно від того, чи всі властивості цих ресурсів сприймаються організмом. Наприклад, істотно важлива якість їжі – її калорійність – не сприймається органами почуттів.
B. До фізіологічного середовища відносяться крім зазначених вище й інші, не необхідні для життя, але все-таки діючі фактори, як правило, значною мірою обумовлюючи оптимальну щільність популяцій й ареали: конкуренти, вороги, паразити, збудники хвороб, несприятливі крайні температури, осмотичний тиск і т.д.
Г. Екологічне середовище включає ще й ті фактори, що діють непрямим чином, впливаючи на мінімальні й фізіологічні фактори. Наприклад, для хижака вигідно, якщо гарне пасовище підвищує щільність популяції його жертв, і невигідно, якщо популяція скорочується через епізоотію або недостачу їжі.
В остаточному підсумку, всі організми й фактори середовища на Землі перебувають у тісному або віддаленому зв'язку між собою. Але тому, що земна поверхня диференційована, виникли більш-менш розмежовані комплекси таких взаємин. Певні групи організмів так зв'язані потоками енергії й речовини, що утворюють досить стабільні у часі й просторі надорганізмові утворення – біоми: ліси, степи, тундри й т.д. Для такого роду екосистем часто характерні певні комбінації видів (співтовариства організмів, біоценози), а також певні комплекси абіотичних факторів середовища.
Безпосереднє оточення організму характеризується з екологічної точки зору пануючими там абіотичними й біотичними факторами. Узяті в сукупності, вони утворюють місцеперебування організму, його «екологічну адресу». Порівняння різних даних про місцеперебування якого-небудь виду дозволяє зробити висновки про фактори, що відносяться до екологічного середовища даного виду й тим самим більш-менш тісно прив'язують його до цих місцеперебувань.
12.2 Умови середовища
12.2.1 Загальні геофізичні умови в біосфері
До загальних умов існування живих організмів відносяться наявність рідкої води, ряду хімічних елементів (так званих біогенних) і надходження променистої енергії в діапазоні температур від –50 до +500С. Тому життя можливе тільки в тонкому (і навіть не скрізь безперервному) шарі між земною корою й атмосферою – у біосфері (рис 12.1).
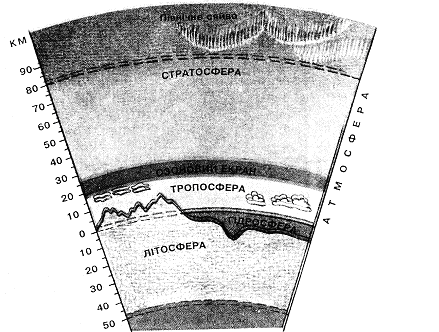
Рис. 12.1 Оболонки Землі
На висотах більше 6000 м та в сухих областях тривалий час здатні виживати тільки спочиваючі стадії (наприклад, спори); у темних глибинах вод і під землею можуть жити лише спеціалізовані консументи й деструєнти, що використовують біомасу, вироблену в освітленій зоні, або біомасу хемосинтезуючих бактерій. В океані освітлена зона рідко заходить глибше 100 м, а в швидкоплинних водах обмежена лише декількома сантиметрами. Хоча біомаса становить по вазі не більше 0,1% земної кори, у неї входять практично всі елементи. Концентрація макроелементів (С, Н, О, N, S, P, Ca, Mg, К, Na, Fe, CI) у біомасі перевищує їхню концентрацію в земній корі, а мікроелементи (наприклад, Al, Zn, Mn, Cu, Si, Br, I, As) виявляються в біомасі звичайно лише в слідових кількостях.
В енергетичному відношенні життя в біосфері підтримується постійним припливом променистої енергії від Сонця й використанням її в процесах фотосинтезу. Цей процес спрямований проти градієнта ентропії, тобто в живих організмах ентропія (що ніби-то протирічить другому закону термодинаміки) зменшується, але це відбувається за рахунок прискореного збільшення ентропії в навколишньому середовищі.
Продуценти (рослини, синьо-зелені водорості, деякі бактерії) здійснюють фотосинтез, при якому відбуваються ендергонічні процеси утворення молекул, збагачених енергією (таких як вуглеводи або білки). Енергія світла перетворюється при цьому в хімічну енергію синтезованих молекул, а пізніше передається гетеротрофним консументам (насамперед тваринам). Зрештою, не витрачена на дихання частина біомаси розкладається деструєнтами, також гетеротрофними (бактерії, гриби й ін.). У підсумку вся біомаса – іноді після тривалої фосилізації – при екзергонічному розпаді вивільняє всю енергію, що міститься в ній. Таким чином, екосистеми, хоча енергія в них на якийсь час затримується, являють собою енергетично відкриті системи. Енергія, що вивільнюється, втрачається для системи (принцип ентропії!), а хімічні елементи можуть використовуватися знову в кругообігу речовин.
Практично всі речовини земної кори з різною швидкістю й у різних кількостях проходять через організми. Існують біогеохімічні кругообіги газового типу з дуже протяжними в просторі й дуже рухливими резервуарами в атмосфері або океанах (цикли О2, СО2, Н2О, N2) (рис. 12.2, 12.3) і кругообіги осадового типу з менш протяжними резервуарами в земній корі (Fe, P, Са).
Крім світла, необхідного для фотосинтезу, вирішальні елементи клімату, особливо для продуцентів, – це температура й вологість. Потік енергії, від якого залежать ці елементи, змінюється від місця до місця (зональні відмінності), протягом доби (добові коливання) і протягом року (сезонні коливання). Ці явища пов'язані з обертанням Землі й нахилом її орбіти до площини обертання. На кліматичні цикли – добові в тропіках і сезонні в помірних поясах, впливає ще й відстань до водних мас океанів (океанічний або континентальний характер клімату).

Рис. 12.2 Кругообіг вуглецю в біосфері
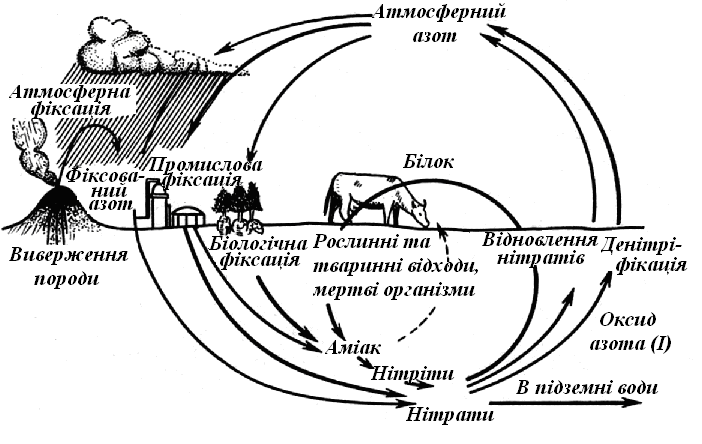
Рис. 12.3 Кругообіг азоту в біосфері
На екоклімат сильно впливає рослинність. При щільному рослинному покриві (поля, лугу, лісу) інтенсивність променевого потоку на його верхній границі така ж, як на голому ґрунті, а визу – часто більш ніж в 10 разів менше. Тому наземна рослинність густого букового лісу асимілює в основному до появи листя на деревах. Оскільки деревний полог гальмує також випромінювання й випар із глибини рослинного покриву, там складається рівний мікроклімат з малими амплітудами коливання температури, підвищеною вологістю повітря й ослабленим його рухом.
Інші розходження в екокліматі визначаються топографією місцевості. Південний схил при сильних розходженнях інсоляції й випромінювання в загальному плані тепліше, сухіше й має більше довгий світловий день, ніж північний схил.
12.2.2 Особливості субстрату
Тому що процеси обміну речовин можуть іти тільки у водяних розчинах, організми спочатку змогли заселити одну лише гідросферу. Пізніше вони вийшли й на сушу в тих місцях, де є досить води, а у вільному повітряному просторі організми можуть перебувати лише тимчасово (при добуванні їжі, розселенні й т.п.).
У більш-менш великих водоймах є два місцеперебування: вільної водної маси і донна зона. І та й інша підрозділяються на нижній, слабко освітлений поверх, де немає фотосинтезу, і верхній, який пронизаний світлом. Сильні плавці (нектон) пересуваються власними зусиллями, а інші – планктони – переносяться плинами. Багато хто із прикріплених або плазуючих мешканців частину життя проводять як планктонні організми.
Для доставки кисню в глибинні шари, а живильних речовин – у верхні вирішальне значення має перемішування води. Для наземних екосистем вирішальне значення має доступність води. Прісна вода становить усього лише близько 3% загального запасу води на Землі (1300 млн. км3). Приблизно 3/4 наявної прісної води лежить у вигляді льоду, а в атмосфері циркулює тільки 0,35%. При цьому 78% води, що випарувалася, циркулює над морем, 14% над сушею й тільки 8% між морем і сушею. Більша частина води атмосферних опадів відразу затримується в рослинності, так що в ґрунт попадає тільки 1/3 –1/4 опадів, що випали. Евапорація (випар з вільної поверхні) і транспірація (випар рослинними тканинами) повертають воду в атмосферу, причому площі, зайняті рослинністю, особливо лісу, віддають більше, ніж голі ділянки. Вода опадів, що не стекла по поверхні в яке-небудь водоймище, просочується у вигляді гравітаційної води в порожнечі ґрунту, де вона в більшому або меншому ступені втримується в капілярах, як плівкова вода (водяна плівка навколо часток ґрунту) або силами когезії як краплинорідинна вода (ґрунтова волога), а частково може проникати в ґрунтові води. Крім ґрунтових вод, у розпорядженні рослин перебуває тільки капілярна вода, що висить у капілярах і тимчасово наповнює порожнечі.
Ґрунт – зовнішній, розпушений фізичним і хімічним вивітрюванням шар земної кори – є одним з найважливіших субстратів життя. Безліч організмів, в основному дрібні (так званий едафон), живе в ґрунті більш-менш постійно. Ці організми вносять вирішальний вклад в утворення ґрунту, механічно розпушуючи й перемішуючи його (дощові черв’яки), сприяючи розчиненню певних речовин (зі звільненням СО2), створюючи запаси відмерлої біомаси й продуктів її розкладу й ремінералізації, у тому числі гумінових речовин. Тому склад і структура ґрунтів – це один з найважливіших факторів, що визначають саме існування як нижчих, так і вищих рослин.
12.3 Організм і середовище
Кожен організм має здатність реагувати у відповідності зі своєю генетичною конституцією на навколишнє середовище, використовувати її фактори для свого існування й розвитку або, принаймні, переносити їхній вплив. Ця екологічна потенція визначається спадкоємною нормою реакції стосовно кожного фактора середовища. Норма реакції у кожного виду характеризується певним положенням і діапазоном (широтою) на шкалі інтенсивності даного фактора – екологічної валентності. Екологічна потенція охоплює діапазон від нижньої песимальної межі (мінімуму) до верхньої песимальної межі (максимуму) з оптимумом десь посередині (рис. 12.4). У песимумах здатність

Рис. 12.4. Екологічна потенція у зіставленні з областю валентності фактора середовища А. Загальна схема для пойкілотермних і гомойотермних тварин. Б. Різні положення й ширина області потенції (норма реакції)
реагувати знижується до нуля, за ними наступає абсолютне гальмування процесу (реакції), часто навіть необоротне ушкодження організму. Широта екологічної потенції (від мінімуму до максимуму) у еврипотентних видів велика, у стенопотентних мала. Дивлячись на те, де лежить оптимум для стенопотентного виду – у нижній, середній або верхній частині всього діапазону, – розрізняють оліго-, мезо- і поліпотентні види. Положення й широта норми реакції залежать від віку, статі й фази розвитку та є різними для різних процесів (активності, метаболізму, розвитку й т.д.). Границі екологічної потенції можуть (зазвичай лише ненабагато) зрушуватися в результаті модифікації, тобто індивідуального пристосування (звикання) або мутації, тобто генетичного пристосування.
12.3.1 Фактор температури
Температура впливає на енергетику всіх життєвих процесів. При підвищенні температури на 10°С реакція прискорюється в 2-3 рази. Однак під впливом ендогенної норми реакції й зовнішніх лімітуючи факторів, відповідна експонентна крива рано або пізно переходить у криву з оптимумом. Активність тварин обмежується песимумами, при яких наступає оборотне теплове або холодове задубіння. Головна точка прикладення для впливу температури – обмін речовин. Для асиміляції, як і для дисиміляції, існують специфічні температурні межі й специфічний оптимум.
Евритермні організми (із широкою екологічною потенцією) – це, наприклад, актинія Actinia equina, що може жити як у тропічних, так й в арктичних морях; коропи, здатні жити й у холодних, і в теплих прісних водоймах. Стенотермні види пристосовані до вузького діапазону температур. Серед них політермні (тобто пристосовані до тепла) майже всі рифоутворюючі корали, терміти, таргани й інші тварини; оліготермі (пристосовані до холоду) гренландський кит, самці зимового п'ядака, струмкова форель, сардинові риби.
Рухливі організми можуть використовувати зовнішні джерела тепла або уникати їх, змінюючи позу стосовно джерела теплового випромінювання (геліорегуляція) або відшукуючи місця, більше підходящі відносно тепла (термотаксис). Термопреферендум (краща температура) специфічний для кожного виду; це та температура, при якій не витрачається енергія ні для виробництва тепла, ні для його віддачі. Організм звичайно знаходить такі умови шляхом фоботаксису, у результаті безладних пересувань (методом проб і помилок). Краща температура, загалом, відповідає екоклімату нормального місцеперебування даного виду. Тільки гомойотермні (теплокровні) тварини – птахи й ссавці – порівняно незалежні від навколишньої температури й створюють свій власний «внутрішній клімат», підтримуючи постійну температуру тіла (37-39°С у ссавців, 41-42°С у птахів). Тому криві їхньої реакції на температуру утворюють майже рівне плато між песимумами (рис. 12.7).
При порівнянні родинних видів з теплих і холодних областей часто можна відзначити збільшення розмірів тіла (правило Бергмана) або зменшення придатків (кінцівок, вух, хвоста; правило Алена) від теплих районів до холодного.
12.3.2. Водний режим
Обмінні процеси – біохімічні реакції, які протікають у водяних розчинах. Тому для активного життя необхідно достатній вміст води в організмі (у наземних тварин 45-95% ваги тіла). Тому що тиск водяної пари в повітрі, як правило, порівняно невеликий, неминучі втрати води в результаті транспірації, а покрити усе тіло водонепроникною ізоляцією (кутикулою, шаром воску, роговим панциром) неможливо через необхідність газообміну при диханні й фотосинтезі. Тому запас води доводиться часто поповнювати шляхом питва (хребетні, павуки, брюхоногі молюски) або споживання вологої їжі (види, що харчуються листами й плодами). Мешканці аридних районів використовують метаболічну воду, що утворюється при окислюванні жирів й інших речовин; жир запасається, наприклад, у горбах верблюдів. При високій вологості повітря можливе поглинання вологи через покриви тіла (амфібії, брюхоногі молюски, тропічні рослини), іноді за рахунок гігроскопічності яких-небудь структур – проти градієнта вологості (мокриці, кліщі, борошняний хрущак). Рослини аридних місцеперебувань мають дуже широко розгалужене або дуже глибоко проникаюче в землю коріння з низьким осмотичним потенціалом.
Втрати на транспірацію зменшуються завдяки захисним покривам і подушкам нерухомого повітря (утримуваного шкірою й пір'ям у тварин, волосками у рослин), зворотному усмоктуванню води при виділенні сечі й дефекації, пошукам вологих місць. Мурахи, терміти, комарі активно розшукують підходящі шари повітря (гігротаксис). Більшості видів властиво фоботаксичне (гігрокінетичне) поводження: їхня ненаправлена активність заспокоюється тільки у місці з підходящою вологістю повітря. Гігрофільні види майже не мають захисту від висихання й можуть існувати тільки в дуже вологому повітрі (амфібії, безраковині брюхоногі молюски, трави тропічного дощового лісу). Ксерофільні види мають або морфологічні засоби захисту від висихання, або можуть накопичувати воду (пустельні сукуленти, наприклад кактуси), всмоктувати її назад, у сухий час доби розшукувати укриття або заповнювати втрати води більшою кількістю вологої їжі (рис. 12.5).
|
|
|
|




Рис. 12.5. Рослини, що належать до різних екологічних груп
12.3.3 Фактор світла
Являючись джерелом енергії для фотосинтезу, світло має фундаментальне екологічне значення. Навіть більш-менш позбавлені світла екосистеми глибинних морських вод, ґрунтів і печер в остаточному підсумку залежать від первинної продукції біомаси зелених рослин на світлі.
L – освітленість у даному місцеперебуванні в порівнянні з повною освітленістю непокритого ґрунту. Світлолюбні рослини пустель, степів, тундр і високогір'я можуть асимілювати тільки при L = 100%. У тіньовитривалих рослин точка компенсації (при якій асиміляція СО2 дорівнює вивільненню СО2) досягається при набагато меншому світловому постачанні, а неослаблене світло їм шкідливе. Лугові рослини займають середнє положення. Світло викликає в органах рослин (стеблах й їхніх розгалуженнях, черешках листів і т.д.) рухи типу скривлень, спрямовані на оптимальне використання світла листами, тобто найвищу активність фотосинтезу.
Так називаним рослинам довгого дня (в основному це рослини середніх широт або що походять відтіля) для початку цвітіння потрібно, загалом, більше тривалий світлий період доби (>12-14 годин на добу), чим рослинам короткого дня, що походять з екваторіальних областей (<12 годин на добу), тому останні можуть у нас цвісти тільки восени (або навесні).
12.3.4. Екологічна потенція
По екологічній потенції відносно температури, світла, вологості повітря, вмісту солей і кисню у воді й т.д. розрізняють види із широкою й вузькою нормою реакції: евритермні і стенотермні, евригідрічні й стеногідричні евригалінні й стеногалінні, евріоксибіонтні й стенооксибіонтні. Лише у деяких видів вимоги до всіх або більшості факторів середовища так малі, а широта реакції відповідно так велика, що вони можуть існувати практично у всіх місцеперебуваннях даної області, тобто бути евріойкними. Повністю евріойкні види й теперішні космополіти рідкісні (людина, пацюк). Стіноойкні види спеціалізовані для життя в певних місцеперебуваннях; але еврипотентні види теж можуть виявитися стеноойкними в тих областях, де їхні вимоги, наприклад до температури, задовольняються тільки в окремих специфічних місцеперебуваннях.
У центрі ареалу даного виду діапазон його потенції часто значною мірою збігається із широтою валентності більшості факторів середовища, а на окраїнах ареалу, навпроти, той або інший фактор часто приходиться на область песимума. Рідко буває так, щоб в одному місці області валентності всі життєво важливі фактори збігалися з діапазонами потенцій виду. Найчастіше хоча б один фактор лежить поза оптимумом. Тоді від цього фактора залежить можливість існування виду в даному місці. Екологічно вирішальним завжди виявляється фактор, що опинився песимальним для тієї стадії, що має найменшу широту реакції.
Вихід за межі песимума не завжди небезпечний для індивідуума. Можливі два способи «емансипації» від екологічної валентності: зрушення діапазону потенції шляхом акліматизації (наприклад, сплячка) і відхилення від несприятливого фактора середовища за допомогою міграції.
Успішність акліматизації до низьких температур (резистентність до холоду) у пойкілотермних (холоднокровних) тварин залежить від попереднього досвіду. У гомойотермних тварин зниження температури діє не тільки як загартування, але й сприяє утворенню підшкірних жирових відкладень і більше густого хутра. У холоднокровних і примітивних теплокровних тварин песимальні умови середовища викликають сплячку, тобто припинення активності й уповільнення процесів метаболізму й розвитку, негайно й неминуче.
Погіршення умов середовища може також приводити до міграцій, причому останні іноді відбуваються завчасно. Регулярні міграції багатьох перелітних птахів визначаються щорічно змінами погоди. Інші («інстинктивні») птахи відлітають у більш-менш фіксовані строки, керуючись, очевидно, фотоперіодом, задовго до погіршення погоди й скорочення харчових ресурсів.
12.4 Популяція й навколишнє середовище
Вирішальні біотичні елементи природних екосистем – це не особини, а популяції. На їхню структуру й динаміку впливають не тільки абіотичні, але, насамперед, біотичні фактори. Щільність популяції (достаток), тобто число (або біомаса) особин на одиницю площі або об’єму, віковий склад і генофонд – основні параметри екології популяцій. Щільність популяції максимальна, коли вона така, що більша щільність уже не могла б підтримуватися даною екосистемою, і мінімальна, коли при меншій щільності члени популяції вже не мали б нормальних шансів на розмноження (на зустріч партнера).
12.4.1. Зміни щільності популяції
Якщо не враховувати імміграцію й еміграцію, щільність популяції визначається внутрішніми (конструктивними) факторами – народжуваністю й смертністю, а на них у свою чергу впливають зовнішні (регуляторні) абіотичні й біотичні фактори.
Загальна народжуваність – це число нових особин, (Nn), що додаються в одиницю часу (t). Тому що воно залежить від числа вже наявних особин, краще використати питому народжуваність:
![]() (12.1)
(12.1)
Ідеальна (максимально можлива) народжуваність реалізується тільки в оптимальних умовах. Опір середовища (недостача їжі, партнерів, місць для розмноження, несприятлива погода й т.п.) зводить ідеальну народжуваність до реальної (екологічної).
Те ж, але зі зворотним знаком вірно й для смертності. Це число особин, що гинуть в одиницю часу (Nm). Питома смертність (d) вийде, якщо віднести цю величину до чисельності популяції:
![]() (12.2)
(12.2)
Ідеальна (мінімальна) смертність проявляється як результат фізіологічної смертності від старості тільки при оптимальних умовах життя. Опір середовища (погода, конкуренція, вороги) підвищує її до реальної (екологічної) смертності.
Від балансу між народжуваністю й смертністю (b d) залежить, якою мірою змінюється щільність популяції. В ідеальних умовах (при максимальній народжуваності, мінімальній смертності й стабільному віковому складі) швидкість росту популяції r1=b-d завжди приймає позитивні значення (b>d), тому що організми завжди дають більше здатного до розмноження потомства, чим було б необхідно, щоб замінити батьків. Тому популяція безупинно росте. Специфічний для кожного виду репродуктивний потенціал (швидкість розмноження, біологічний потенціал, або потенційна сила розмноження)
![]() (12.3)
(12.3)
при повній відсутності опору середовища дає експонентний ріст популяції.
У природних умовах ріст популяції рано або пізно припиняється через опір середовища, що збільшується вже через те, що зростає щільність популяції. Тому реальна крива росту звичайно приймає сигмовидну (логістичну) форму.
Розмір популяції підтримується на рівні К різними способами. У видів, що живуть у ефемерних місцеперебуваннях з високим опором середовища (більші втрати від ворогів), або в паразитів (малі шанси знайти хазяїна) репродуктивний потенціал п повинен бути дуже більшим (r1-стратеги), щоб вони могли швидко використати рідкі або невеликі шанси. Навпроти, види, що живуть у довгострокових, стабільних місцеперебуваннях, з невеликою кількістю ворогів або зовсім їх не мають, види з розвитий турботою про потомство, що утворять сім'ї або стада, обходяться невеликим репродуктивним потенціалом, тому що в них значна частина нащадків досягає репродуктивного віку. Наприклад, орли, дельфіни й великі копитні дають у рік лише одного нащадка.
12.4.2 Вплив біологічних факторів
У регулюванні щільності популяцій відіграють значну роль внутрішньовидові й міжвидові взаємодії. Особливо велика роль конкуренції за ресурси. Внутрішньовидова конкуренція (за їжу, статевого партнера, життєвий простір, місце для розмноження) збільшується з ростом щільності популяції й ступеня спеціалізації виду. Найчастіше починається конкуренція за їжу, коли в результаті розмноження при ще достатньому запасі їжі щільність популяції підвищується. Недостатнє харчування може тоді приводити, наприклад, до зниження плідності, поки зменшення популяції не дозволить виду знову розмножуватися.
Навіть при достатній кількості їжі висока щільність – фактор скупченості – може знижувати плідність. Висока щільність іноді приводить до канібалізму навіть у видів у нормі чисто рослиноїдних.
Організми протидіють занадто високої щільності популяції за так називаним принципом конкурентного виключення. Активна протидія приймає форму територіального поводження, пасивна складається з дисперсії (розсіювання), тобто розрідженні популяції шляхом більше рівномірного розподілу в просторі.
Територіальність – явище, засноване на вродженому прагненні особини до волі пересування на деякій мінімальній площі. Перший щабель розвитку територіальності – індивідуальний простір, що оточує кожну особину (воно добре помітно в ластівок, що сидять на телефонних проводах, або у шпаків у зграї, що летить). Особина захищає його від вторгнення й відкриває для іншої особини тільки після церемоній залицяння перед спарюванням. Другий щабель – оборона місце для життя, відпочинку або сну у середині не оборонної зони активності (у багатьох хижаків – мисливської ділянки). Саме раціональне використання простору досягається на третьому щаблі територіальності, коли утворюються справжні території – ділянки, з яких інші особини виганяють.
Міжвидові взаємини можуть бути байдужними, шкідливими або корисними для партнерів. При нейтралізмі обидва види живуть в одній екосистемі, не вступаючи у відносини один з одним (наприклад, гидроїдні поліпи та раковині молюска, дятли неподалік від дроздів у буковому лісі). Може існувати конкуренція за однакову їжу або життєвий простір (наприклад, між коріннями дерев у савані, між польовими горобцями й синицями за місця гніздування). Мутуалізм приносить вигоду обом партнерам – при симбіозі життєво важливу, при протокооперації – не дуже значну (рис. 12.6). Жуйні тварини й мікроорганізми їх рубця не можуть існувати один без одного; навпроти, гідра може жити без водорості хлорели, як і та без її.
Частіше користь і шкода бувають однобічними. Для лева байдуже, чи поїдають грифи й шакали залишки його трапези (коменсалізм) (рис. 12.7); для жуків-гнойовиків несуттєво, що в польоті вони переносять нематод-копрофагів до нових гнойових куп – їхньому субстрату (форезія). При паразитизмі й хижацтві один з партнерів витягає для себе користь на шкоду іншому. Ці два типи взаємин розрізняються тим, що в першому випадку нападаючий організм менше своєї жертви, а в другому – крупніше. Крім того, репродуктивний потенціал у паразита більше, ніж у хазяїна, а у хижака — менше, ніж у жертви.
|
|
| |
|
Актинія на панцирі раку пустельника |
Лишайник | |
|
|
| |
|
Симбіоз орхідеї й гриба. А – квітки північної орхідеї-ятришника. Б – гіфи гриба в клітинах ятришника, В – проросток орхідеї з ділянкою, зайнятою міцелієм |
Схема зв'язку міцелію гриба з коріннями дерева – мікоризи | |
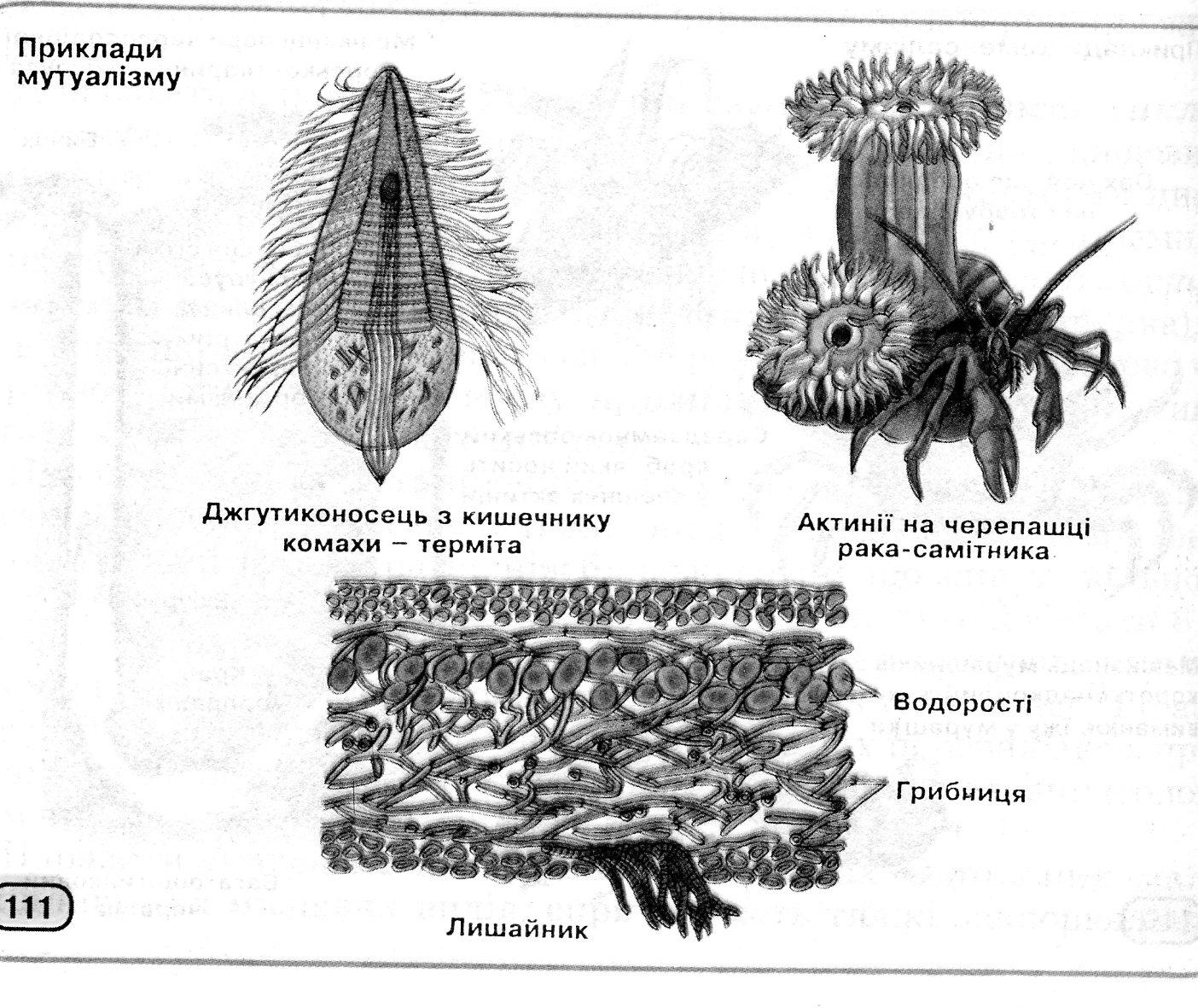
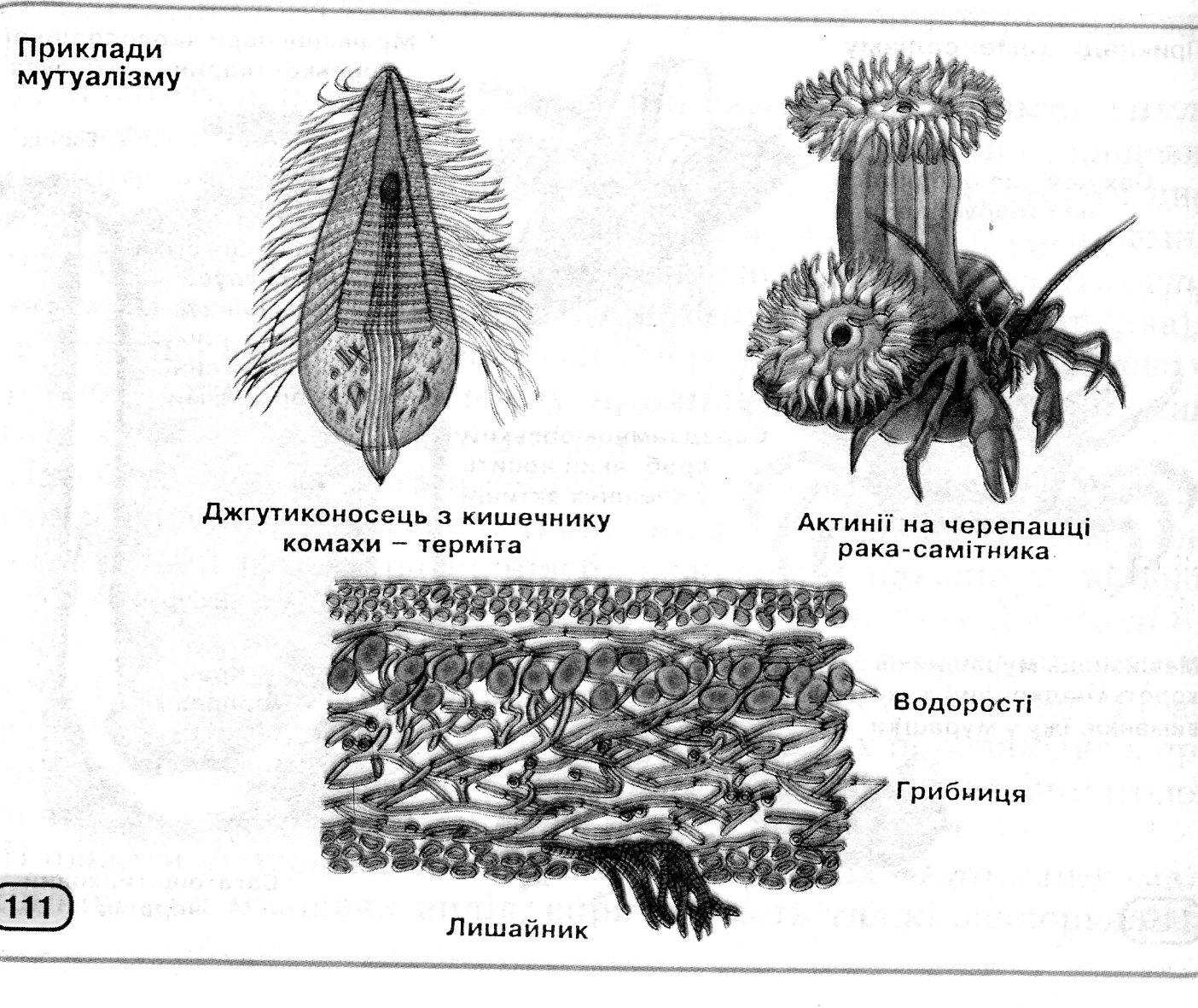


Рис. 12.6 Симбіоз
|
|
|
|
Орхідея, яка оселилась на стовбурі дерева |
Середньоморський краб, який носить у клешнях актинії |
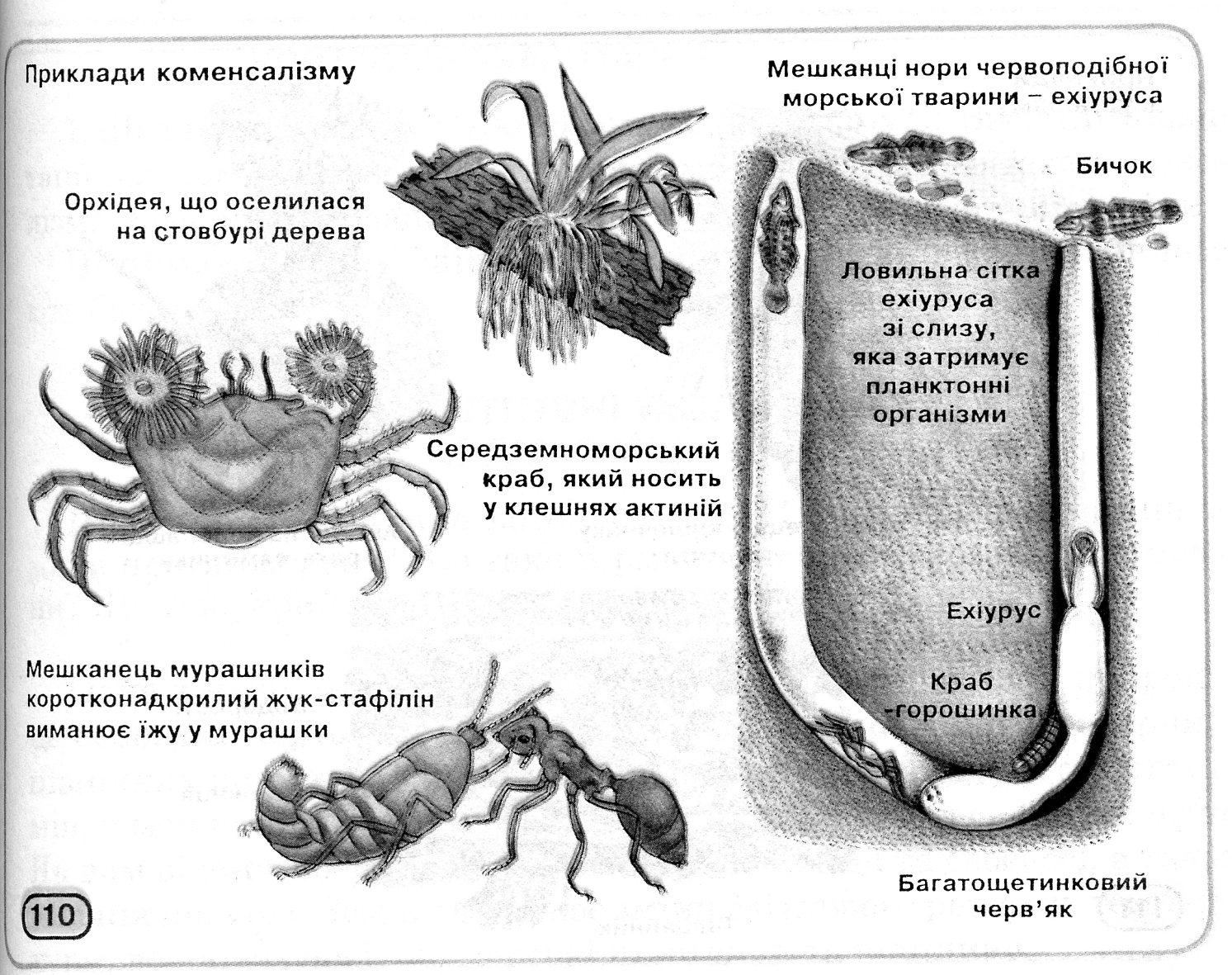
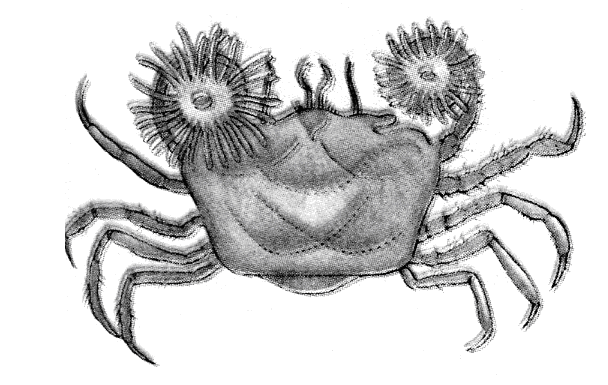
Рис. 12.7. Приклади коменсалізму
У гомогенному середовищі, що не має укриттів для розмноження, хижак рано або пізно знищує популяцію жертви й після цього вимирає сам. У природних умовах виникає наступний тимчасовий й причинно – слідчий ланцюг: розмноження жертви → розмноження хижака → різке скорочення чисельності жертви → падіння чисельності хижака → розмноження жертви й т.д. Ця кібернетична система з негативним зворотним зв'язком приводить до стійкої рівноваги; хвилі флуктуації хижака й жертви випливають одна за одною з постійним зрушенням по фазі, і в середньому чисельність як хижака, так і жертви залишається постійною.
Для популяції жертви вплив хижака позитивний, тому що надмірне розмноження жертви привело б знову до різкого падіння її чисельності. До того ж хижак в основному винищує хворих і слабких особин, які діють як фертильні внутрішньовидові конкуренти. У свою чергу всі механізми, що перешкоджають повному винищуванню жертви, сприяють збереженню харчової бази хижака.
Захист від ворогів може бути активним – наприклад, укуси, уколи, удари (у тому числі електричні – в електричних скатів, сомів й інших риб), вибризкування секретів і т.п., використання укриттів – або, набагато частіше, пасивним, до якого відносяться ті, що маскують (мімікрійна) зовнішність; зовнішність, що застерігає (мімікрія в широкому змісті слова); поведінка, що маскує або застерігає. У рослин розвиваються колючки, шипи, пекучі волоски, отрути, гіркі речовини.
Зовнішність що маскує, полягає в наслідуванні неїстівним предметам, (наприклад, палочники й гусениці п'ядаків імітують сучки, паличник «бродячий лист» і відпочиваючі метелики – листи, довгоносик Llthinus – лишайники) або зоровому злитті з навколишнім тлом: зелене фарбування мешканців листя (квакші, коники, клопи, гусениці), коричнева – у наземних мешканців (жайворонки, пісочники, самки качок). Іноді пристосування до кольору й візерунку субстрату може здійснюватися шляхом фізіологічної зміни фарбування тіла (каракатиці, скати, камбали, квакші) або зміною фарбування при черговому линянні (коники).
Застережлива зовнішність може використатися із двома цілями. Одна мета – відлякування агресора незвичайним малюнком, очними плямами, що з'являються в багатьох метеликів, коли вони розкривають крила, імітацією зміїної голови (у багатьох гусениць) або імітацією тварин, небезпечних для нападників (відлякувальна зовнішність). Інша можливість – попередження яскравими сигнальними кольорами й малюнком, що кидаються в очі, про реальні негативні для нападаючих властивостях жертви: гіркому смаку, неїстівності, отруйності, умінні кусати або жалити. Приклади – сонечка, клопи-арлекіни, строкаті гусениці, оси. Але при цьому приноситься в жертву якась частина популяції, на якій агресор засвоює гіркий досвід. Деякі нешкідливі організми імітують попереджуюче фарбування небезпечних видів (наприклад, мухи-дзюрчалки, багато жуків-вусанів і метелики-бражники наслідують зовнішності ос). Вони, можна сказати, користуються захистом своїх зразків, копіюючи їх: ця мімікрія у вузькому змісті слова – обман.
12.4.3 Регулювання щільності популяції
Ріст популяції регулюється факторами залежними і не залежними від її щільності.
Не залежні від щільності фактори, такі як більшість кліматичних факторів (температура, вологість повітря, кількість опадів й ін.), прискорюють або гальмують ріст популяції завжди в ступені, пропорційному вихідній щільності. Те ж можна сказати й про сприятливі погодні умови.
На відміну від цього фактори, що залежать від щільності, як, наприклад, більшість біотичних факторів (конкуренція, вороги, паразити, патогени), впливають на ріст популяції тільки лімітуючим чином і їхня дія ніколи не буває однаковою при різній щільності популяції. Вони діють як регулятори у вузькому змісті слова, «підганяючи» популяцію до існуючій у цей момент ємності середовища. Окремий хижак скорочує щільну популяцію жертв менше, ніж рідку, але тільки в тому випадку, якщо він не розмножується. Таким чином, дія факторів, що залежать від щільності, сильно залежить також від загальної ситуації в екосистемі.
12.5 Екосистеми
Екосистеми - це надорганізмові об'єднання з певним видовим складом. В екосистемах суші (лісах, степах, лугах, болотах і т.д.), насамперед, впадають в око нерухомі, звичайно добре помітні продуценти, що мають велику біомасу. Склад рослинних співтовариств в екосистемах характеризується не систематичними категоріями, а життєвими формами. Переходи між екосистемами часто поступові (наприклад, між лісом і степом), границі їх нечіткі (наприклад, між очеретяним болотом й озером). Будучи енергетично й структурно відкритими системами, вони зобов'язані своєю єдністю взаємодії своїх компонентів, які підтримують у відношенні один до одного й до неживого середовища статистичну (рухливу) рівновагу. Тому екосистеми характеризуються не стільки певними числами й нерухомими границями, скільки середніми значеннями й границями розкиду.
12.5.1 Структура екосистем
З багатьох подібних за своєю біологією видів однієї екосистеми (наприклад, комах, що смокчуть соки лугових рослин, – попелиць, цикад, клопів) звичайно лише деякі (5-10%) становлять основну частину (близько 4/5) біомаси даної групи. Висока щільність популяцій цих домінуючих видів говорить про їхню оптимальну пристосованість до даної екосистемі й великому для неї значенні. В однакових екосистемах, віддалених одна від одної областей, один вид може бути замінений іншим: наприклад, в американських тропічних лісах серед птахів запильниками квіток служать колібрі, а в африканських – медососи.
Для виду як структурного елемента екосистеми вирішальне значення має не його систематичне положення, а функціональна конституція й екологічна потенція. Тому вид являє собою комплекс життєвих форм, які дозволяють йому використовувати певні властивості (екологічні ліцензії) середовища (субстрати, джерела їжі, діапазони температур і вологості й т.д.) і грати в такий спосіб певну роль в екосистемі.
Як один вид може утворювати цілий спектр життєвих форм, так кожне місцеперебування надає організму цілий спектр факторів у вигляді екологічних ліцензій для використання. І з того і іншого – зі сполучення області потенції певної життєвої форми й області валентності необхідних для її реалізації факторів середовища – при достатньому збігу складається екологічна ніша. Життєву форму можна зрівняти із професією, наявну валентність – з робочим місцем, а екологічну нішу – з реалізованими трудовими відносинами.
Можливість впровадження випадково завезених (колорадський жук) або мимовільно поширених (кільчаста горлиця) видів показує, що в екосистемі – по історичним й екофауністичним або екофлористичним причинам – ще можуть бути вільні, довго не використовувані ліцензії.
Сукупність всіх видів, які зустрічаються спільно, називають співтовариством організмів, або біоценозом, а зайняте ними місцеперебування, тобто сукупність всіх екологічних факторів їхнього місцезнаходження, – біотопом («місце життя»). Але оскільки біоценоз важко чітко відокремити від біотопу, краще поєднувати їх під ім'ям біогеоценозу.
Більшість біогеоценозів підрозділяється на субодиниці (яруси, біохори, меротопи. Найбільш стабільні й замкнуті яруси («поверхи»), наприклад, у лісі деревинний ярус, чагарниковий, трав'янистий і надґрунтовий. Хоча кожний з них має власний мікроклімат, а часто й власну фауну (тлі, короїди), вони зв'язані між собою кліматично й функціонально (наприклад, харчовими ланцюгами за участю листового опаду).
У кожну екосистему входять наступні функціональні компоненти:
а) продуценти – автотрофні організми, які з використанням сонячної енергії будують із неорганічних сполук багату енергією біомасу (зелені рослини, синьо-зелені водорості, фото- і хемосинтезуючі бактерії);
б) консументи – гетеротрофні організми, які використовують цей органічний матеріал для одержання й нагромадження енергії, змінюють або перебудовують органічні речовини (тваринні й гетеротрофні мікроорганізми);
в) деструєнти – гетеротрофи, які руйнують використані або відмерлі залишки біомаси, розкладають їх на неорганічні складові частини (мінералізація), що надходять знову в резервуари мінеральних речовин (бактерії, гриби).
Консументи харчуються живим (біофаги) або мертвим (сапрофаги) органічним матеріалом. Серед біофагів можуть бути виділені рослиноїдні організми, або фітофаги (первинні консументи, до них відносяться також віруси, що вражають рослини, бактерії, гриби й паразитичні судинні рослини), хижаки (вторинні консументи, у їхньому числі й паразити первинних консументів) і кінцеві споживачі – верхові хижаки (третинні консументи).
Представники розглянутих вище трофічних категорій (трофічних рівнів) зв'язані між собою односторонньо спрямованою передачею біомаси у харчові ланцюги (ланцюга харчування). Всі харчові ланцюги входять у кругообіги матерії, що ведуть від продуцентів до деструєнтів; ланцюги можуть складатися з малої або великої кількості ланок.
Пасовищні харчові ланцюги (рис. 12.8), або ланцюги хижаків, починаються із продуцентів: конюшина – вівця – вовк; планктонні водорості – дафнія – плотва – щука – скопи. Для таких харчових ланцюгів характерне збільшення розмірів особин (висхідні ланцюги) при одночасному зменшенні щільності популяцій, швидкості розмноження й продуктивності по біомасі. Ланцюги паразитів можуть починатися із продуцентів (яблуня – щитівка – наїзник) або консументів (вівця – муха-кровососка – бактерії – віруси); для них характерне підвищення щільності популяції й швидкості розмноження при зменшувані розмірів особин.
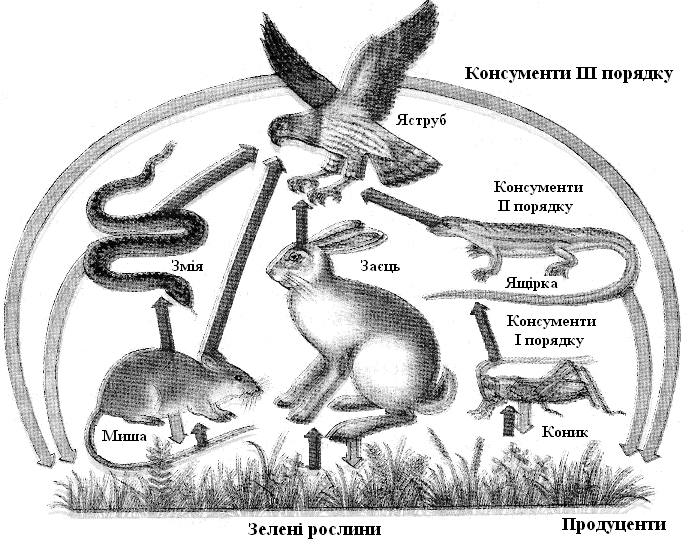
Рис. 12.8. Трофічні ланцюги в наземному біогеоценозі
Детритні ланцюги проходять через відмерлу біомасу; вони звичайно короткі (листовий опад – дощові черв’яки – бактерії).
Тому що продуцентів звичайно поїдають різні консументи, хижаки використовують багато видів жертв, а всеїдні організми (людина, ведмідь, горобець) споживають як продуцентів, так і консументів, тобто живуть на різних трофічних рівнях, харчові ланцюги багаторазово розгалужуються й сплітаються в складні харчові мережі.
Якщо розташувати один над одним графічні відображення щільності популяцій (число особин на 1 м2), біомаси (у грамах сухої або вологої ваги на 1 м2) або продуктивності в енергетичних еквівалентах (джоулях на 1 м2 у рік) для всіх членів кожного трофічного рівня (в одному харчовому ланцюзі), вийде так називана харчова (екологічна) піраміда (рис. 12.9). піраміди енергії (тому що в них ураховується час) стоять завжди «правильно». Навпроти, «догори ногами» стоять піраміди чисел для ланцюгів паразитів і ланцюгів, що починаються з деревоподібних продуцентів.
12.5.2 Фізіологія екосистем
Екосистема – відкрита система, у якій процеси утворення й подальшої передачі біомаси йдуть за рахунок припливу енергії сонячного випромінювання. При кожній передачі на наступний трофічний рівень частина доступної енергії не сприймається, частина віддається (у вигляді тепла), а частина витрачається на дихання. За грубою оцінкою, при цьому загальна енергія щораз зменшується в десять разів, а при передачі від первинних продуцентів консументам – навіть у сто разів. Наслідок цього – обмежена довжина харчових ланцюгів, а також зменшення кількості особин і загальної енергії при переході від нижчих рівнів до вищого. Чим більше довжина харчовий ланцюг, тим менше доступної енергії залишається до її кінця.
12.5.3 Розвиток екосистеми
Хоча екосистеми, перебуваючи в стаціонарному стані, відрізняються статистичною сталістю своїх елементів, вони можуть розвиватися, переходячи від більше простих до більше складних форм.
Там, де відступаюче море, озеро, що висихає, льодовик або діяльність людини звільняють незайману сушу, а також там, де з вулканічного попелу, з лави або в результаті вивітрювання гірських порід утворюється новий ґрунт, починається її заселення рідкою, одноярусною піонерною рослинністю. З нагромадженням гумусу й пилу, з підвищенням вологості ґрунтів розвиваються двох'ярусні вторинні співтовариства, луги й степи. Стійка кінцева стадія (клімакс) цієї первинної сукцесії – у більшості випадків багатоярус ний ліс (рис. 12.16).

Рис. 12.9. Екологічні піраміди.. Коники поїдають зелені рослини, жаби – коників, змії – жаб, орел – змій.
Вторинні сукцесії як би заліковують ушкодження, природні або нанесені людиною (наслідку бурі, вирубки, пожежі, повені, випасу худоби). Яким буде кінцевий етап сукцесії, залежить насамперед від макроклімату (кліматичний клімакс). Наприклад, на більших висотах й у високих широтах клімаксним співтовариством буде вже не ліс, а тундра. На сукцесію впливають також едафічні (ґрунтові, водні) і топографічні особливості (ухил, експозиція, рівень ґрунтових вод). Так само може впливати діяльність людини – випас худоби, косовиця, рільництво (параклімакс), так що ландшафти найчастіше являють собою мозаїку з екосистем з різним ступенем зрілості.
Сукцесії екосистем, як й еволюція живого на всіх рівнях, спрямовані на забезпечення подальшого існування, на гомеостаз. Очевидно, що стабільність рухливої рівноваги при мінливих умовах середовища легше всього досягається в тому випадку, якщо екосистема складається з максимально можливого числа компонентів (внутрішня розмаїтість); тоді екологічні потенції різних видів можуть так доповнювати один одного, що різні перешкоди, як зовнішні (особливо непередбачені зміни абіотичних факторів, до яких неможливо пристосуватися), так і внутрішні (надмірні репродуктивні потенціали деяких організмів), будуть згладжуватися.
12.6 Людина й навколишнє середовище
Людина, будучи всеїдним консументом, спочатку займала порівняно скромне місце в деяких стабільних екосистемах. Маючи видатний духовний потенціал, винаходячи й виготовляючи знаряддя й зброю, вона, однак, усе більше звільнялась від своїх природних екологічних зв'язків і сама ставала найважливішим екологічним фактором, з об'єкта еволюції перетворюючись в її суб'єкт. Оволодіння вогнем і розвиток землеробства й скотарства дозволили їй відволікати на себе енергетичні потоки інших екосистем. Так вона навчилася використовувати високу продуктивність ранньої фази в сукцесіях природних екосистем по можливості необмежено в просторі й часі. Для цього вона стала спочатку шляхом корчування, випалювання, меліорації або зрошення зводити підходящі клімаксні співтовариства до початкових стадій, а потім зробила ці стадії постійними, підтримуючи монокультури, знімаючи майже весь урожай біомаси й щорічно обновляючи ґрунти оранкою.
Більш-менш паралельно із цими подіями людина довідалась про фазу легкої запам'ятованості стимулів-об'єктів у ранній молодості в стадних тварин. Цю особливість удалося використати для їхнього приручення, одержавши слухняних свійських тварин. Проміжки між урожаями, обумовлені зміною пір року, зробили необхідним появу економіки, заснованої на запасах, і звідси, видимо, виникла віра, що не залишає нас дотепер, у необхідність постійного зростання виробництва. Так була запущена та фатальна спіраль розвитку людства, у якій виробництво надлишків неминуче веде до подальшого експонентного росту населення, а цей ріст, у свою чергу, робить необхідним подальше розширення виробництва.
Це прагнення до максимізації й неприродне, і неекологічне, тому що однобічний розвиток одного компонента згодом руйнує будь-яку систему. Воно змушує безперервно боротися з опором середовища, що спочатку успішно протидіяло експонентному росту людства і його виробництва. Хоча в потенції кожен організм здатний до експонентного розмноження, у природних екосистемах воно раніше або пізніше перетвориться під тиском протидіючих біотичних й абіотичних факторів (погодних явищ, ворогів, паразитів, збудників хвороб і т.д.) у логістичний ріст; у результаті між всіма членами біоценозу встановлюється рівновага, що забезпечує стабільність біоценозу. Це неминучий процес, що не вимагає припливу енергії ззовні.
Але, якщо сукцесія штучно зупиняється на лабільній початковій стадії й при цьому неухильно підвищується продукція біомаси (що постійно вилучається) одного-єдиного виду – культурної рослини або свійської тварини, – доводиться не тільки відшкодовувати мінеральні речовини, що витрачаються, за допомогою добрив, але й вносити додаткову енергію, щоб усувати протидіючі види («бур'яни», «шкідники»), які використовуюють монокультуру для себе, змушують її до логістичного росту або прагнуть довести сукцесію до клімаксу. При цьому в принципі байдуже, чи застосовуються проти таких ворогів хімічні (отрути), біологічні (інші, «корисні», організми) або змішані засоби боротьби. У кожному разі необхідне детальне знання біології й екологічних потенцій всіх компонентів системи, включаючи їхню здатність до опору шляхом модифікаційної або мутаційної адаптації.
Безпосередні вороги самої людини – хижаки й збудники хвороб (паразити й патогени) – у результаті полювання й досягнень медицини (у тому числі гігієни) настільки відтиснуті, що хижаки в усім світі практично вже не мають для людини значення, а паразити у високорозвинених міських цивілізаціях грають лише підлеглу роль. Патогенні мікроорганізми – малюсінькі n-стратеги, що мають величезний потенціал розмноження, який полегшує їм можливість утворення штамів, резистентних до лікарських речовин, і нових вірулентних штамів, – усе ще викликають небезпечні епідемії. І все-таки середня тривалість життя людини виросла з 18 (у бронзове століття) до 70 років (у Європі й Північній Америці). Це значно сприяло демографічному вибуху XX століття, у який, втім, особливо значний внесок внесли країни Азії й Південної Америки, що розвиваються,. На підвищення щільності населення (різне в різних регіонах) накладається всі зростаюча урбанізація. Близько 1800 р. у містах з населенням 20 000 чоловік і більше жило менш 5% усього людства, в 1950 р. – 21%, а зараз – 30%. На початку ХІХ століття на Землі було 27 великих міст із населенням 100 000 чоловік і більше, а наприкінці ХХ століття – більше 1800 таких міст.
Дотепер людина одержувала необхідну їй додаткову енергію майже винятково з викопних енергетичних запасів, що нагромадилися у формі вугілля, нафти й газу в результаті утворення біомаси в минулі епохи. Але їхнє вичерпання в доступному для огляду майбутньому неминуче, і тому необхідно шукати інші джерела енергії. Тільки завдяки копальним пальним матеріалам стали взагалі можливими процеси індустріалізації, які побічно сприяли розгортанню небезпечної спіралі росту населення. У результаті навколишнім середовищем для людства стала тепер практично вся біосфера, але для підтримки панування над нею людині потрібно усе більше енергії. Хоча на Землю продовжують надходити величезні кількості сонячної енергії, жоден технічний процес не зміг поки задовільно відтворити (а тим більше перевершити) унікальну процедуру використання цієї енергії у фотосинтезі зелених рослин. До того ж нам погрожує і вичерпання непоновлюваних матеріальних ресурсів, наприклад запасів урану, срібла, цинку.
Людина активно використовує у своєму сільському й лісовому господарстві майже всі наземні біоми. Але зараз її діяльність зачіпає й практично всі інші природні екосистеми – на них впливають хоча б промислові відходи, для яких у біосфері немає деструєнтів (наприклад, пластмаси). Ці екосистеми вже змінені (ДДТ в організмі антарктичних пінгвінів і в льоді Арктики) або ушкоджені (забруднення океанів нафтою, а атмосфери – SО2) антропогенними впливами й далеко відхилилися від стаціонарного стану. Тому усе наполегливіше висувається вимога створити економіку, нешкідливу для природи, з обмеженим ростом при оптимальній чисельності населення. Але ще не відомо, чи допускає взагалі генетичний потенціал людини таку якісну зміну життєвих потреб, пов'язану із самообмеженням і скороченням споживання, тим більше що людина вже в такій значній мері звільнилася від дії сил природного добору. Доти поки ідеологія зростання виробництва навіть охорону природи розглядає лише як засіб до «кращого використання ресурсів» і прагне в усім світі не до оптимуму, а до максимуму, людство не підійде до «екологічної моралі», що сприяла б розвитку екологічної економіки, тобто розвитку цивілізації не за рахунок природи й проти її, а в стійкій гармонії з нею.
Великі перспективи відкриває контроль над народжуваністю, що уже сьогодні привів в індустріальних суспільствах до стабілізації чисельності населення. З підвищенням рівня життя в країнах, що розвиваються, таке положення може, нарешті, стати загальною нормою. Але тоді постійний ріст майна й влади, ресурсів й енергії перестане бути мірою добробуту. Адже такий світогляд уже зробило людину єдиною істотою, що налагодила організоване винищування собі подібних. Таким чином, загальну боротьбу проти війни можна розглядати як багатообіцяючий підхід до людської екології – до створення самими людьми навколишнього середовища, більш гідного людини.
Запитання для самоперевірки
Як можна розділити «шари» навколишнього середовище за характером відносин між організмом та елементами середовища?
Загальні геофізичні умови в біосфері. Потік енергії через екосистеми.
Біогеохімічні круговороти карбону та азоту.
Клімат.
Організм й навколишнє середовище Екологічна потенція.
Фактори температури, водного режиму та сітла.
Популяція й навколишнє середовище. Щільності популяції
Вплив на популяцію біологічних факторів. Мутуалізм, хижацтво та паразитизм.
Захист від ворогів.
Регулювання щільності популяції
Екосистеми: структура та функціональні компоненти.
Трофічні категорії (трофічні рівні). Харчові ланцюги, мережі, піраміди.
Розвиток екосистеми.
Особливості антропогенного впливу на природне середовище.
Основні напрямки збереження природного середовища Землі.