

Глава 11 еволюція
План
11.1 Сутність еволюції
11.2 Фактори еволюції
11.3 Шляхи еволюції
11.1 Сутність еволюції
Кожен окремий організм розвивається (індивідуальний розвиток, онтогенез). Крім того, у довгому ряді поколінь від предкової форми до нащадків виникають спадкоємні зміни, що призводять до нових видів, зміненим планам будови й типам функціонування, причому складність організації зазвичай зростає. Цей ряд змін називають філогенезом, або еволюцією.
Умови, що підходять для життя, створилися на Землі лише на певному етапі її історії, і тоді живе повинне було розвитися з неживого. У джерел біологічного розвитку стояли дрібні, дуже просто влаштовані агрегати. Із самих пристосованих серед них через безперервний ланцюг поколінь розвинулося все існуюче наразі різноманіття організмів. Еволюційний розвиток не повертає назад, це історично необоротний процес.
Спадкоємні зміни є необхідними для філогенетичного розвитку й разом із саморепродукцією і дискретністю індивідуумів відносяться до основних ознак живого. У взаємодії з добором (і іншими факторами) вони уможливлюють філогенетичний прогрес, тобто еволюцію, у напрямку оптимального функціонування в існуючих умовах.
Цілями еволюційної теорії повинні бути:
доказ існування філогенетичних змін;
за можливістю повне з'ясування шляхів філогенетичної зміни в різних групах організмів, тобто побудова філогенетичних дерев (дендрограм) і систем, що відбивають природне споріднення;
з'ясування причин і принципів дії, які лежать в основі рушійних сил еволюції.
11.1.1 Докази еволюції
Важливий матеріал, що доводить існування філогенетичних змін, дала порівняльна морфологія, яка виявила спільність плану будови в споріднених організмах і дозволила охарактеризувати основні типи цього плану, а з іншого боку – виявила зміну функцій морфологічних структур. При цьому вирішальне значення для філогенетичних побудов набув принцип гомології, а аналогічні (конвергентні) структури, навпроти, не могли служити підставою для висновків щодо споріднення (рис. 11.1). Гомологічними називають структури загального філогенетичного походження (коли риси подібності обумовлені спорідненням), а аналогічними – функціонально подібні, але різні за походженням структури (коли риси подібності обумовлені пристосуванням до однакового середовища) (рис.11.2).

Рис. 11.1. Конвергенція. Розвиток органів ширяння у ширяючих хребетних.
А – летюча риба; Б – літаюча жаба; В – літаюча агама; Г – білка-летяга.
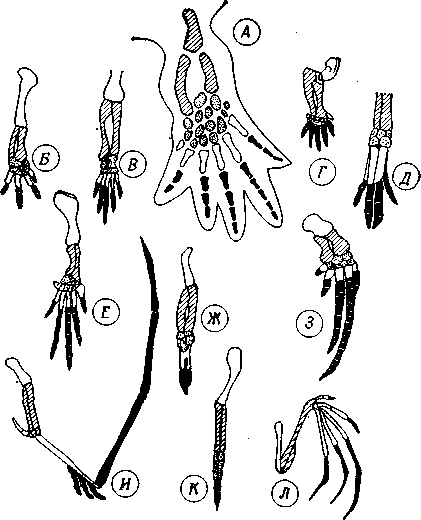
Рис. 11.2. Гомологічні органи. Гомологія кінцівок чотириногих, встановлена на основі гомотопії. А. Основний план будови кінцівки чотириногого. Б-Д. Тварини із земноводним або наземним способом життя: Б – саламандра, В – крокодил, Г – кріт, Д – свиня. Е-З. Плаваючі тварини: Е – морська черепаха, Ж – пінгвін, 3 – кит. И-Л. Літаючі тварини: И – літаючий ящір, К – птах, Л – кажан
Інші свідчення на користь еволюції, теж засновані, в остаточному підсумку, на принципі гомології, можна знайти в процесах ембріонального розвитку, як це сформулював Е.Гекель у своєму біогенетичному законі (рис. 11.3): онтогенез у скороченому виді повторює деякі фази, пройдені даною групою організмів у філогенезі. Крім того, цінні відомості може дати наявність рудиментарних (залишкових, недорозвинених) органів або поява атавізмів (повернення до вже втрачених ознак), що нагадують ознаки предкової форми (рис. 11.4).
|
1 |
|
|
2 | |
|
3 | |
|
|
А Б В Г Д Е Ж З |
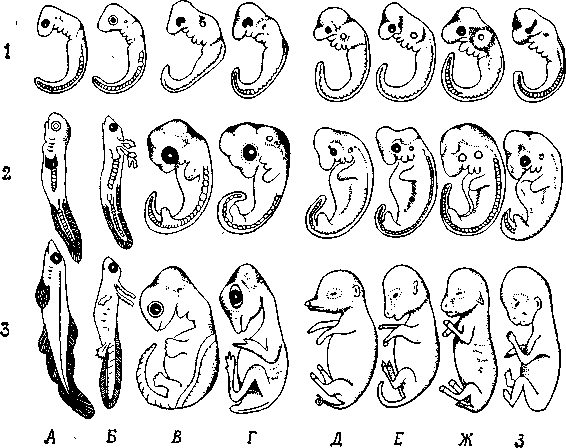
Рис. 11.4. Подібність ембріональних стадій як прояв «біогенетичного закону». 1, 2, 3 – стадії розвитку. А – риба; Б – саламандра; В – черепаха; Г – птах; Д – свиня; Е – корова; Ж – кролик; З – людина. (Наеckel.)
|
|
|
| |||
|
Третє віко Вушна раковина |
|
Сліпа кишка с червоподібним відростком | |||
|
|
|
|
| ||
|
|
Волосата людина |
Багатососковість у людини |
Хвостатий хлопчик | ||
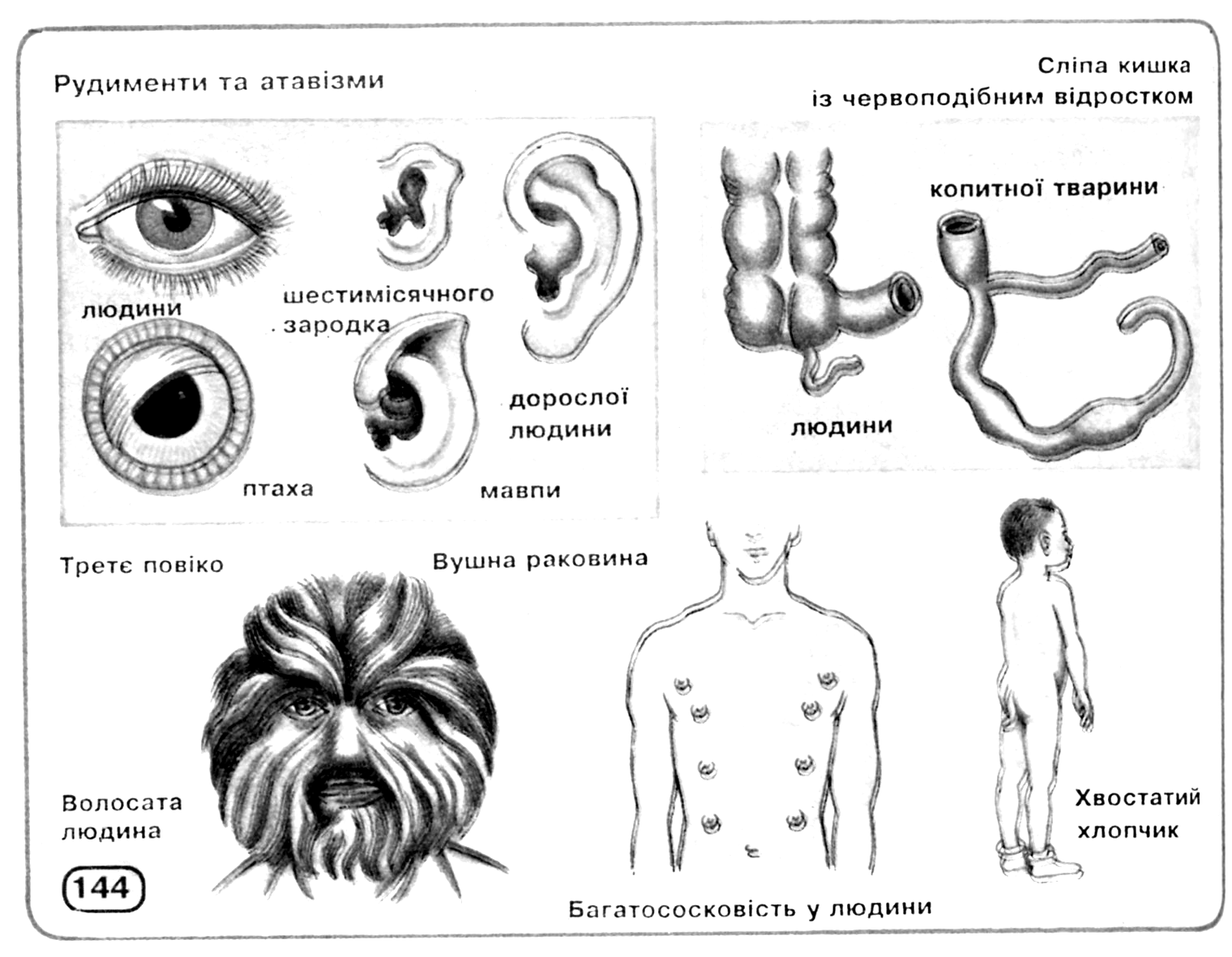
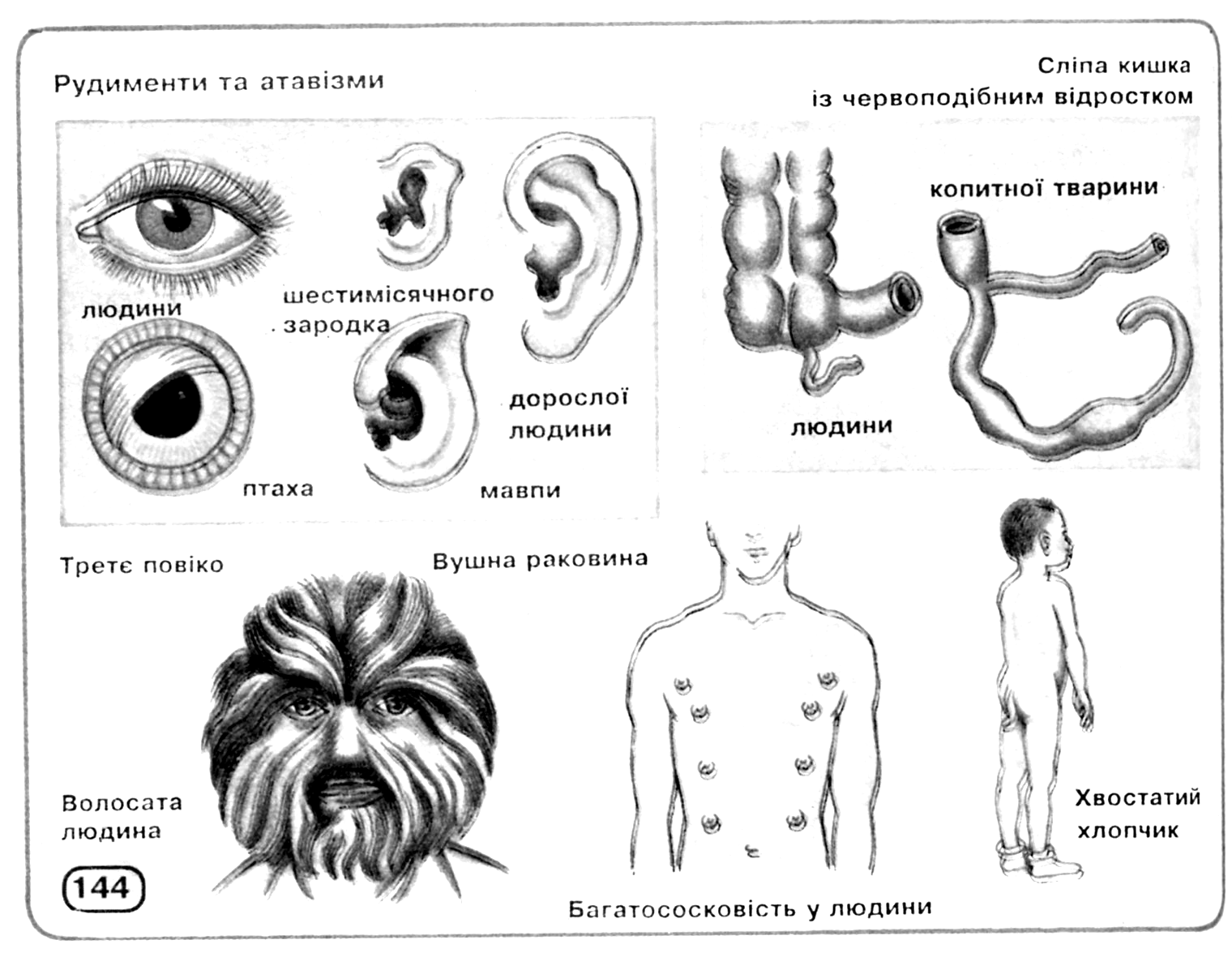
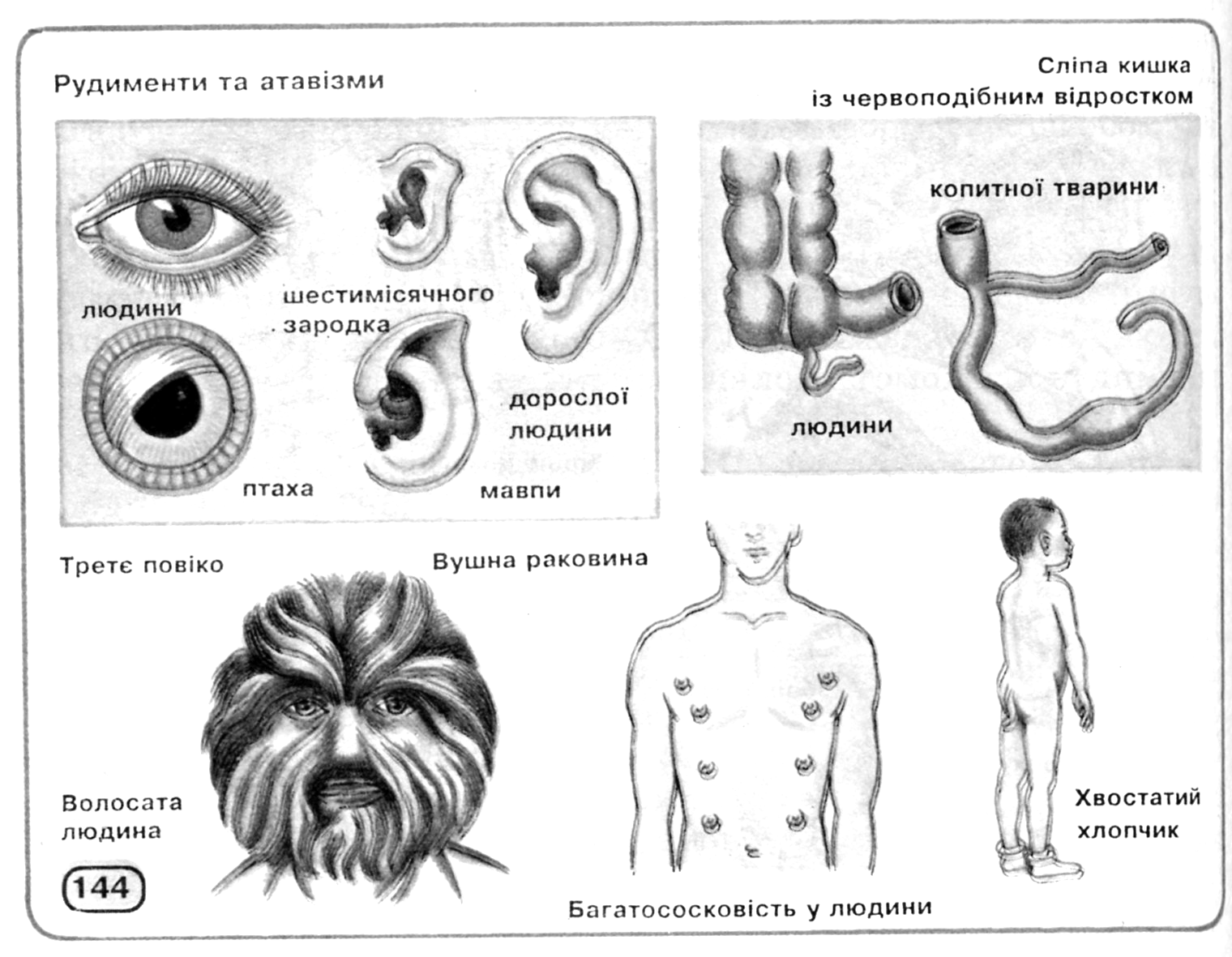
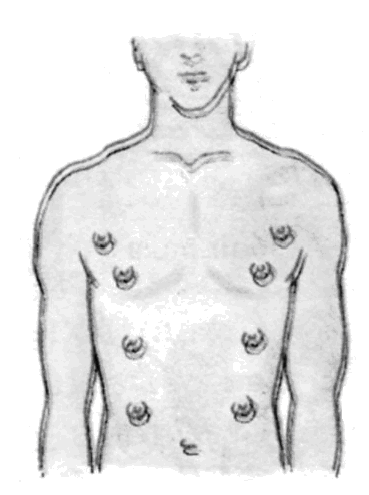
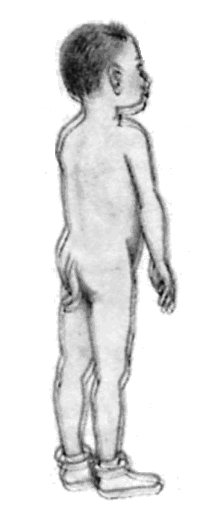
Рис. 11.4. Рудименти і атавізми
Палеонтологія накопичила безліч викопних знахідок, а також даних щодо умов життя в колишні епохи. Молекулярна (хімічна) палеонтологія займається хімічним складом ранніх слідів життя з відкладень, вік яких може перевищувати 3,4 млрд. років.
Нарешті, біогеографія і екологія теж доставляють факти, важливі для розуміння еволюції. Так, ендемічне (просторово обмежене) поширення, наприклад, однопрохідних ссавців або гатерій свідчить про їхній реліктовий характер. Вивчення екології паразитів і їхньої приуроченості до хазяїв допомагає виявити родинні взаємини.
11.1.2 Еволюційні теорії
Історія еволюційних теорій характеризується зміною різних уявлень щодо чинників, які сприяли доцільній адаптації організмів до навколишнього середовища. Ж.-Б.Ламарк висував на перший план спрямовані пристосування, обумовлені прямим впливом середовища і можливі завдяки «прагненню організмів до вдосконалення». Він вважав, що придбані ознаки успадковуються (ламаркізм). Це подання, так само як і думка про прямий вплив навколишнього середовища на еволюційні події, виявилося помилковим.
Ч.Дарвін і незалежно від нього А.Уоллес обґрунтували принцип природного добору й уявлення про «боротьбу за існування» як механізмі цього добору (теорія природного добору, дарвінізм).
Дарвін висував наступні положення:
а) близько родинні організми (наприклад, батьки й діти) подібні між собою, але в них є спадкоємні розходження;
б) ці розходження стають більше помітними, якщо розглядати довгі ряди предків і нащадків;
в) різні ознаки змінюються з різною швидкістю, так що одні ознаки можуть бути більш давнім філогенетичним придбанням, ніж інші;
г) виробляється більше нащадків, ніж може вижити, і тому відбувається добір «найбільш пристосованих».
Таким чином було визнано, що еволюція обумовлена природними причинами і це відкрило шлях до їхнього наукового аналізу.
Подальший розвиток ідей Дарвіна, насамперед роботами Хакслі і Симпсона, призвело до «синтетичної теорії еволюції». Ця теорія пояснює різноманіття і пристосованість організмів як результат дії в основному двох факторів: безперервної появи спадкоємних відхилень і добору, що здійснюється факторами зовнішнього середовища. Онтогенез і фізіологічні процеси розглядаються як реалізація спадкоємної інформації, а філогенез – як формування й випробування усе нових і нових спадкоємних інформаційних програм.
11.2 Фактори еволюції
11.2.1 Вид і його визначення
У центрі уявлень Ч.Дарвіна щодо еволюції є поняття виду. До Ч.Дарвіна вид вважали незмінним, і ця незмінність служила вирішальним аргументом для будь-якого антиеволюціоністського напряму думок. Існування еволюції можна було довести тільки продемонструвавши, що один вид може виникнути від іншого шляхом спадкоємної зміни.
Для виділення «виду» використовують в основному два підходи: один з них ґрунтується на морфологічних ознаках, інший – на особливостях біології розмноження й екології.
Таксономист має справу в основному з фіксованим, а палеонтолог – тільки з викопним матеріалом, тому вони у своїй роботі можуть опиратися майже виключно на морфологічні ознаки. Із цього погляду вид визначають як групу особин, найбільш подібних між собою за важливими для систематики морфологічним ознакам, – виділений за цим критерієм вид називають морфологічним видом.
Однак поряд із цим усе ширше використовуються також фізіологічні й біохімічні ознаки, важливі для життєдіяльності. З біологічних критеріїв особливу роль грає схрещуваність представників одного виду. Можливість розмноження усередині виду забезпечується подібністю генетичного матеріалу, тобто збігом числа і структури хромосом і наявністю видоспецифічних генів. Таким чином, вид являє собою сукупність подібних і здатних до схрещування між собою індивідуумів, у репродуктивному відношенні ізольовану від інших подібних сукупностей. Окремі співтовариства, що розмножуються в собі, усередині виду та мають загальний генофонд, називають популяціями.
Види розділені репродуктивними бар'єрами – особливостями, що запобігають схрещуванню. Це розходження у формах поводження, несумісність гамет, стерильність батьків або гібридних особин (в останніх не утворюються функціонуючі гамети або статеві органи).
В екологічному відношенні для особин одного виду характерні однакові взаємини із середовищем: кожен вид займає свою особливу екологічну нішу. Розмежування за особливостями біології розмноження й екологічних ознак приводить до біологічного уявлення щодо виду, до біологічного виду.
Таким чином, вид – єдина реально існуюча категорія. Його загальний генофонд забезпечує достатню мінливість, але, з іншого боку, настільки єдиний, що може підтримувати досить стабільний гомеостаз виду як співтовариства, що розмножується, і екологічної єдності.
Еволюційні зміни в межах виду називають внутрішньовидовою еволюцією; їм протиставляють зміни, що виходять за межі виду – надвидова еволюція. В основі внутрішньовидової і надвидової еволюції, безсумнівно, лежать ті ж самі механізми.
11.2.2 Виникнення спадкоємних варіантів
Особини зі спадкоємними змінами виникають у результаті мутацій і схрещувань (рекомбінації) організмів з різними алелями або генами. Поки особливості розмноження, морфології або екології змінені лише незначно, нові популяції залишаються в складі вихідного виду. Тільки після явного виникнення механізмів ізоляції, що відокремлюють їх від вихідного виду, можна говорити про новий вид (або більш високому таксоні).
У результаті генних мутацій виникають нові алели, які приводять до несприятливих, сприятливих або байдужних змін вихідного типу. Мутанти з несприятливими змінами більш-менш швидко елімінуються, якщо це гаплоїдні організми або диплоїди з домінантною мутацією. При рецесивній мутації новий алель може зберегтися в популяції у гетерозиготному стані. Алель, що у гомозиготному стані викликає несприятливий ефект (як, наприклад, ген серпоподібноклітинної анемії), у гетерозиготному стані іноді може створювати селективну перевагу (у даному прикладі він знижує сприйнятливість до малярії). У нейтральних мутантів спадкоємна зміна спочатку не проявляється у фенотипі. Ґрунтуючись на змінених таки чином амінокислотних послідовностях, вдавалося будувати еволюційні схеми й робити висновки щодо родинних зв'язків між організмами.
Филогенетичне дерево, засноване на даних про зміни в гені rРНК 16S (малої субчастки рибосом) свідчить про походження прокаріот й еукаріот від загальних предків.
Ступінь подібності виражали як відношення
![]() ,
,
де А и В – порівнювані види. Величині SAb відповідає на схемі положення самої нижньої горизонтальної лінії, що з'єднує обидва види. При SAb = 1,0 збіг був би повним.
Про значення сприятливих мутацій часто можна судити за порівняно швидким поширенням їхніх носіїв, особливо у тому випадку, якщо вони добре пристосовані до умов, що змінилися. У березового п'ядака (Biston betularia) крім світлої нормальної форми зустрічається темний мутант (саrbonaria). Ці дві форми розрізняються алелями одного гена і у разі схрещування дають менделівське розщеплення. В Англії у областях із сильним задимленням атмосфери переважає мутантна форма, а в районах з чистішим повітрям – нормальна, тобто в кожному випадку та, котра менш помітна для ворогів.
Одна або кілька генних мутацій, як правило, приводять лише до внутрішньовидової еволюції. Границі виду долаються тільки у результаті численних (або особливо важливих) мутацій, якщо виникаюча при цьому нова форма ізолюється від вихідного виду репродуктивними бар'єрами.
У процесі еволюції відбувалися великі зміни числа хромосом. У сучасних організмів на один геном доводиться від 1 до 500 і більше хромосом. Їх число часто буває поліплоїдним або анеуплоїдним. Поліпоїди становлять 30-50% всіх квіткових рослин (це в більшості випадків алополіпоїди). Після поліплоїдізації в результаті неправильного розходження хромосом може відбутися анеуплоідизація.
У ході еволюції відбувся величезний ріст кількості ДНК у геномі: у бактерій один геном містить 106-107 пар нуклеотидів, у ссавців – близько 5-109. Однак у вищих організмів близько 30% ДНК повторюється, у бактерій же повторюваної ДНК усього лише близько 0,3%.
Схрещування – набагато більш часті події, ніж мутації, особливо у видів з перехресним заплідненням. Приводячи до рекомбінації генів, вони значно підвищують різноманіття форм усередині виду.
11.2.3 Напрямні фактори
Вихідним матеріалом для всякої еволюції служать генетичні варіанти. Генетичні зміни самі по собі позбавлені доцільності, вони відбуваються у всіляких напрямках. Зіштовхуючись із різноманітними внутрішніми (морфологічними і функціональними) і зовнішніми (навколишнє середовище) умовами, змінені особини одержують різні шанси передати потомству свою змінену генетичну інформацію і цим вплинути на частоту певних алелей у популяції. Цей принцип називають принципом добору; це статистичний процес. Слова Дарвіна щодо добору як «виживанні найбільш пристосованих» і про «боротьбу за існування» нерідко розумілися невірно – як «боротьба всіх проти всіх».
Селективна цінність (ступінь адаптивності, пристосованість) генотипу визначається його реакцією на тиск добору, тобто суму всіх діючих селективних факторів. Ступінь їхнього впливу вимірюють коефіцієнтом добору s. При повній елімінації (потомства немає) s=1; якщо ж частота алеля не зменшується (у добре пристосованих форм), s=0. Процеси добору піддаються математичному аналізу й моделюванню.
Добір, впливаючи в основному на особини, робить у генному фонді спрямовану зміну, що у ряді поколінь (гологенії) викликає підвищення пристосованості. Цю можливість спрямованого добору використовує і людина у формі штучного добору в тваринництві та рослинництві; вона заснована на разючій мінливості видів (рис. 11.5).
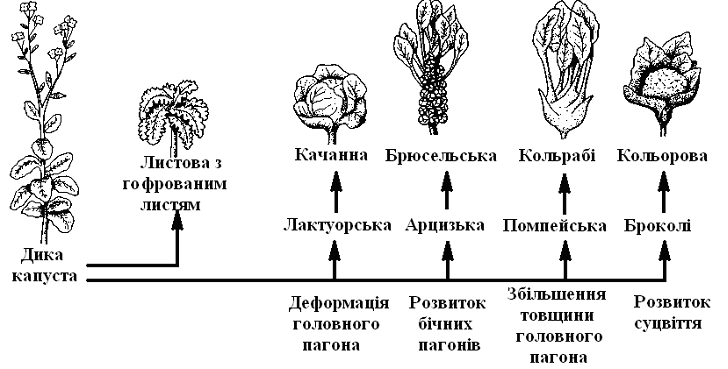
Рис. 11.5 Штучний добір: результати селекції капусти (Rundfeld)
Добір забезпечує збереження ознак, що виправдали себе, і в цьому сенсі він консервативний (стабілізуючий добір). З іншого боку, він заохочує усі нові форми із вдалими сполученнями ознак і тому прогресивний (трансформуючий або динамічний добір).
Ізоляція частин популяції створює бар'єри для схрещування; порушується безперешкодний обмін генами з батьківською групою – панміксію усередині виду. Тому ізоляція – важлива передумова видоутворення.
Географічна ізоляція може створюватися як результат активного або пасивного розселення, зміни клімату (наприклад, у епохи заледеніння), геоморфологічних змін (утворення островів, гороутворення) або в результаті впровадження в ареал непридатних для заселення просторів (пустель, водойм). Все це веде до так називаного алопатрічного видоутворення, при якому виникаючі види відособлені в просторі.
Генетична ізоляція може спричиняти утворення нових видів і без просторового поділу. Таке видоутворення називається симпатричним. Інші механізми ізоляції: екологічне відокремлення, тобто використання різних екологічних ніш в одній і тій же області поширення, фізіологічні розходження, морфологічна дивергенція, відхилення поведінкових ознак, розходження у геномі.
У ході еволюції досягають успіху ті організми, які краще інших пристосовані до навколишнього середовища. Якщо умови середовища зміняться, то організм, щоб вижити й продовжувати розмножуватися, може адаптуватися шляхом модифікації, але тільки в рамках спадкоємно певної для нього норми реакції.
Поряд із цими модифікаційними пристосуваннями існують і спадкоємні адаптації, що виходять за межі норми реакції. На системах, зручних для генетичного аналізу, було показано, що це обумовлено наявністю мутацій, що поліпшують пристосованість: чисельність відповідних мутантів через їхню селективну перевагу зростає.
11.2.4 Еволюція на надвидових рівнях
Кожен план будови як деяка загальна ознака пристосований до відповідного середовища. Якщо добір протягом тривалого часу діє в одному напрямку (ортоселекция), то створюється стійка, порівняно прямолінійна еволюційна тенденція, і це дозволяє пояснити згадані вище випадки «спрямованої» еволюції (ортогенезу).
Вироблення нових типів організації – типогенез – відбувається шляхом невеликих кроків, результати яких підсумовуються (адитивний типогенез) і можуть підсилюватися завдяки взаємодії субсистем (органів). Поступовість цих змін підтверджується рядами викопних форм. Носії ознак предкового типу й нового типу зв'язані перехідними формами, так званими «проміжними ланками».
Будь-яка зміна умов середовища – скажімо, кормової бази тощо – змінює тиск добору (наприклад, убік утворення корінних зубів з високою коронкою у травоїдних тварин). З іншого боку, структури, що звільняються від тиску добору (наприклад, фоторецептори у печерних тварин або ендопаразитів), перетворюються у рудиментарні (залишкові) органи і, зрештою, можуть зовсім зникнути заради економії.
Коли даний тип організації опанує нову екологічну зону, починається пристосування до специфічних біотопів цієї зони. Зберігаючи свої основні риси, вихідний тип як «загальна» форма спеціалізується в різних напрямках. Еволюційні лінії ніби то розходяться променями, тому говорять про адаптивну радіацію. Цей процес обумовлений «відцентровою силою» добору.
Дарвінові в'юрки (Geospicinae) Галапагоських островів, що представлені більш ніж 10 видами, походять від одного виду, що був занесений на острови наприкінці третинного періоду з південноамериканського континенту. Очевидно, вони виявилися тут першими сухопутними птахами і знайшли місцеперебування, вільне від конкурентів. Адаптивна радіація, що почалася, привела до поділу виду на форми, – тих, що харчуються на землі, і мешканців мангрового пояса, густих лісів і відкритих просторів. У зв'язку з конкуренцією за їжу відбулася спеціалізація у харчуванні: ми знаходимо тут зерноїдні види з конусоподібним дзьобом, комахоїдні види з довгими тонкими дзьобами й навіть дятлових в'юрків, які здатні своїм сильним дзьобом розкривати ходи комах у деревині, але через відсутність довгого язику витягають видобуток за допомогою гострих гілочок й інших подібних знарядь (рис. 11.6).
Значення дивергенції, що прямо наводить на думку про еволюційний процес, і конвергенції, що вказує на механізм еволюційних змін, особливо добре видні на прикладі паралельної еволюції сумчастих і плацентарних ссавців. Обидві групи перетерпіли конвергентну еволюцію і у результаті зайняли ідентичні екологічні ніші в різних частинах земної кулі (рис. 11.7 і табл. 11.1).
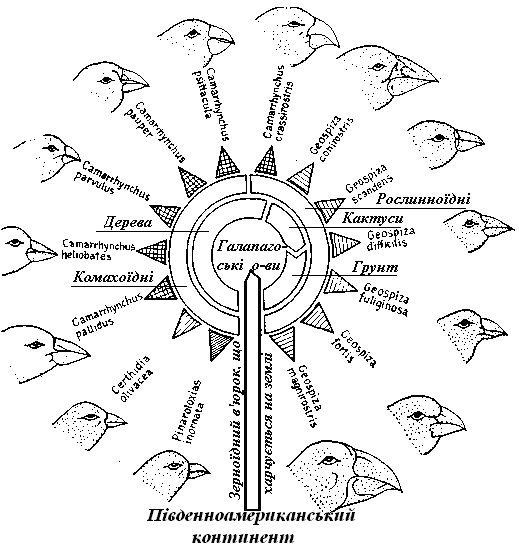
Рис. 11.6. Адаптивна радіація: Дарвінові в'юрки на Галапагоських островах
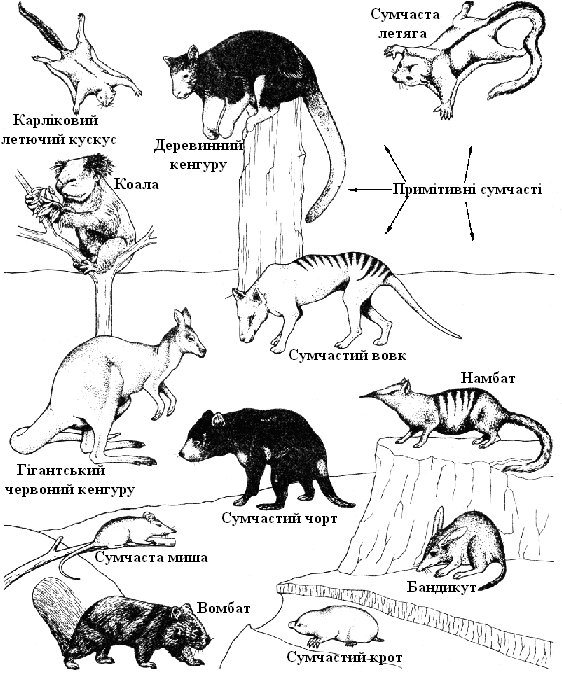
Рис. 11.7. Сумчасті в Австралії
Таблиця 11.1