1. Імунітет 2. Механізми реакції антиген – антитіло 3. Тромбоцити 4. Протизгортальна система крові 5. Кровотворення і його регуляція
1. Імунітет Традиційно під поняттям імунітету розуміли несприйнятливість багатоклітинного організму (макроорганізму) до інфекційних захворювань. Ця властивість забезпечується багатьма системами живого макроорганізму. Наприклад, шкіра, епітелій дихальних шляхів, слизова оболонка травного каналу за їх механічної цілості є непроникними для мікроорганізмів. Захисну функцію виконують також хімічно активні середовища – хлоридна (соляна) кислота шлункового соку, нормальна мікрофлора кишок. Проте головну роль у захисті макроорганізму від інфекції відіграє система крові.
Ще в минулому столітті було відкрито явище фагоцитозу і створено клітинну теорію імунітету (І. Мечников). Тоді ж було виявлено протимікробні властивості плазми крові, що дало початокгуморальній теорії імунітету (П. Ерліх). Сучасна імунологія визнає рівноправне існування обох механізмів імунітету. Кожен з них може бути як специфічним, так і неспецифічним.
Неспецифічний імунітет властивий тваринам усіх рівнів розвитку і, в свою чергу, поділяється на спадковий і клітинний (фагоцитарний). В основі спадкового імунітету лежать загальнобіологічні явища спадковості, мінливості та природного добору. Якщо в популяції макроорганізмів, яка потерпає від певної інфекції, в результаті випадкової мутації з'являється індивід, білкові молекули якого не асимілюються патогенним мікроорганізмом-агресором або є токсичними для нього, то цей індивід чи група індивідів виживає і дає життєздатне потомство, а решта популяції гине. Щоправда, мікроорганізми також модифікуються і пристосовуються до нових умов, з часом захоплюючи й цей ареал, тобто процес боротьби за виживання між макро- і патогенними мікроорганізмами відбувається постійно. Тому є підстави вважати, що мікроорганізми та їхні молекулярні патогенні чинники є важливими і чи не єдиними біотичними агентами, що продовжують природний добір серед людей.
До чинників спадкового імунітету, що розвинулись у процесі еволюції, належать протимікробні та противірусні чинники.
Лізоцим (мурамідаза) – білок, що руйнує бактеріальні мембрани. Він міститься в білку курячого яйця, у слині, сльозах, у складі кишкового соку, скелетних м'язах, мозку, а також у гранулах нейтрофільних гранулоцитів. Крім бактеріолітичної дії лізоцим стимулює також синтез антитіл.
Комплемент – термочутливий (інактивується нагріванням) комплекс з більш ніж 20 білків, здатних до самоорганізації в систему. Більшість білків цієї системи перебувають у плазмі крові в неактивному стані у вигляді проферментів, які активуються в певній послідовності у разі контакту з бактеріями й вірусами. Активація комплементу спричинює бактеріолізис, стимулює фагоцитоз, продукцію та виділення тканинами речовин, які беруть участь у запальних процесах. В активації комплементу бере участь білок сироватки крові пропердин, який має виражену протибактеріальну та противірусну активність.
Інтерферон– це низькомолекулярний білок, що продукується лейкоцитами, діє на клітини, інфіковані вірусом, не прямо, а стимулюючи вироблення противірусних речовин сусідніми неінфікованими клітинами макроорганізму.
Катіонні білки – дефензинита гістони – виявляють високу протибактеріальну та противірусну активність.
Більшість зазначених чинників виробляються нейтрофільними та еозинофільними гранулоцитами, моноцитами і діють у самій клітині, в її фагосомах або виділяються в кров і виконують свої функції поблизу від лейкоцитів. Усі вони є елементами системи неспецифічного спадкового гуморального імунітету.
До неспецифічного імунітету належить також клітинний імунітет, зумовлений активністю лейкоцитів і тромбоцитів. Найбільшу фагоцитарну активність виявляють нейтрофільні гранулоцити і моноцити. Зокрема, частина моноцитів, потрапляючи в тканини, перетворюється там намакрофаги. Для цих клітин не має значення вид мікроорганізму чи природа токсину. Якщо вони здатні знищити це чужорідне тіло, то захоплюють його і перетравлюють. Нещодавно було відкрито різновид лімфоцитів – NK-лімфоцити (природні убивці – naturalkillers), здатних за допомогою білкаперфорину без попередньої сенсибілізації знищувати пухлинні та інфіковані вірусами клітини макроорганізму. Ці NK-лімфоцити являють собою «першу лінію оборони» макроорганізму, оскільки вони реагують на появу чужорідних клітин негайно.
Специфічний (гуморальний, лімфоїдний) імунітет, властивий лише хребетним тваринам, здійснюється лімфоцитами. Лімфоцити, що виходять у кров із кісткового мозку, імунологічно є нейтральними, або нульовими. Імунокомпетентними вони стають після дозрівання або диференціації в органах лімфоїдної системи (мал. 11).
Мал. 11
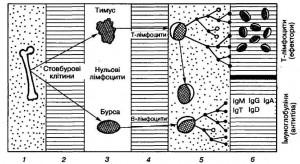
Специфічний імунітет 1– кістковий мозок; 2, 4, 5, 6 – кров; 3 – центральні лімфоїдні органи
Частина нульових лімфоцитів з кров'ю потрапляє до загруднинної залози – тимуса і внаслідок складної диференціації перетворюється на імунокомпетентні Т-лімфоцити, тобто такі, що здатні вступати в реакцію з антигеном (бактеріями, чужорідним білком тощо). Вперше зустрівши певний антиген, Т-лімфоцит «запам'ятовує» його і починає ділитись (проліферувати). Більша частина новоутворених Т-лімфоцитів вступає в реакцію з антигеном і за допомогою перфорину знищує його. Це Т-лімфоцити-кілери (вбивці). Частина Т-лімфоцитів у реакцію не вступає і продовжує циркулювати з кров'ю, іноді все життя. Це лімфоцити імунологічної пам'яті. У разі повторного контакту їх з таким самим антигеном вони «впізнають» його, починають інтенсивно ділитись, утворюючи велику кількість Т-лімфоцитів-убивць, які й знищують антиген. Такого типу реакцію називають вторинною імунною відповіддю, і її перебіг значно інтенсивніший, ніж під час першої зустрічі з антигеном (первинна відповідь).
Частина клітин імунологічної пам'яті продовжує циркулювати в організмі до наступного контакту з антигеном. Зрозуміло, що різні Т-лімфоцити «запам'ятовують» і налаштовуються на реакцію з різними антигенами, але кожний лімфоцит розпізнає лише один антиген. У цьому і полягає специфічність такого імунітету, хоча сам діючий чинник Т-кілерів – перфорин – є неспецифічним. Серед Т-лімфоцитів розрізняють також Т-хелпери (помічники), без яких лімфоцити-кілери не можуть виконувати свою функцію, лімфоцити-супресори, які пригнічують імунні реакції, та інші.
Друга частина нульових лімфоцитів проходить диференціацію в лімфатичних вузлах кишок, апендикса та кістковому мозку. Вони дістали назву В-лімфоцитів, оскільки вперше цей процес було досліджено у птахів, у яких він відбувається у сумці (bursa) Фабриція. Новоутворені молоді лімфоцити потоком крові розносяться до лімфоїдних утворів різних органів, де і відбувається диференціація, внаслідок якої вони стають імунекомпетентними, але ще не є зрілими ефекторними В-лімфоцитами. На їхній поверхні містяться вже готові молекули імуноглобуліну – антитіла до конкретного антигену. При першому контакті В-лімфоцита з антигеном здійснюється «запам'ятовування» антигену і проліферація В-лімфоцитів. Більша частина дочірніх клітин осідає в центрах розмноження в лімфоїдній системі організму і перетворюється на плазматичні клітини, що продукують антитіла, – виникає первинна гуморальна імунна відповідь.
Решта В-лімфоцитів знову виходять у кров і стають лімфоцитами імунологічної пам'яті. У разі появи антигену В-лімфоцити починають синтезувати антитіла до того антигену, що спровокував цю імунну реакцію, які, пройшовши крізь їхню мембрану, переходять у тканинну рідину та кров. І вже за межами В-лімфоцитів, у плазмі чи в тканинах відбувається реакція антиген – антитіло, внаслідок якої антиген знешкоджується під час наступних імунних реакцій. Це вторинна гуморальна імунна відповідь, яка відбувається значно активніше і швидше, ніж первинна, а також швидше (десятки хвилин, години), ніж вторинна клітинна імунна відповідь, спричинена Т-лімфоцитами (1-2 доби). Відповідно першу з них називають імунною реакцією негайного, а другу – сповільненого типу.
Виходячи з описаних вище особливостей реагування імунної системи на антиген, у медичній практиці використовують засіб специфічної профілактики інфекційних хвороб – вакцинацію. Вона полягає в тому, що попередньо здійснюється штучний контакт макроорганізму з ослабленим інфекційним агентом, який не спричинює захворювання, але зумовлює появу лімфоцитів імунологічної пам'яті до цього антигену. У разі повторного, вже не спровокованого контакту макроорганізму з цим антигеном лімфоцити проліферують і здійснюють ефективну імунну реакцію, запобігаючи захворюванню.
Т- і В-лімфоцити різняться не тільки за походженням і властивостями, а й за механізмом дії. Так, рецептори Т-лімфоцитів відрізняють «своє» від «чужого» або зміненого «свого» завдяки наявності на поверхні клітин антигенів гістосумісності і специфічно реагують на певний антиген, але здійснюють імунну реакцію за допомогою неспецифічного чинника перфорину чи інших протеаз. Що стосується В-лімфоцитів, то розміщені на їхній мембрані рецептори і є антитілами, що продукуються В-лімфоцитами під час імунної реакції й безпосередньо вступають у специфічну реакцію з відповідним антигеном за межами лімфоцита.
Обидві групи лімфоцитів досить тісно взаємодіють між собою. Зокрема, Т-лімфоцити-хелпери можуть активізувати синтез антитіл В-лімфоцитами, а Т-супресори, навпаки, пригнічують гуморальні реакції антиген – антитіло, зумовлені В-лімфоцитами. Існує також чітка взаємодія між системами, що забезпечують специфічний і неспецифічний імунітет. Так, неспецифічний чинник – комплемент – за наявності специфічних антитіл спричинює лізис бактерій. Макрофаги передають антигенну інформацію про перетравлені ними мікроорганізми Т-лімфоцитам-кілерам.
Механізми реакції антиген-антитіло
Антиген – це органічна речовина біологічного походження, здатна викликати імунну реакцію. Антигеном може бути шкідлива чи навіть нешкідлива для організму речовина, але, як правило, чужорідна для нього. В антигені можна виділити дві частини – молекулу-носія, імунологічно нейтральну високомолекулярну органічну сполуку, та розміщені на її поверхні кілька низькомолекулярних груп – гаптенів, які мають органічну чи неорганічну природу і надають усій молекулі здатності спричинювати імунну відповідь. Такі антигени називають повними на відміну віднеповних антигенів, до яких належать речовини, зокрема гаптени, що самі не здатні викликати імунну відповідь, але набувають такої здатності після сполучення з білком-носієм. Так, зокрема, низькомолекулярні речовини, потрапивши до організму, можуть набувати антигенних властивостей після їх сполучення з макромолекулами цього організму. В такому разі вони виступають у ролі гаптенів.
Антигени є видові, а у хребетних тварин також групові та індивідуальні, тканинні тощо. Така висока специфічність антигенів визначається певними хімічними групами – детермінантами специфічності, або епітопами, завдяки яким антиген розпізнається антитілом. Детермінанта специфічності – це невелика частина молекули антигену (4-10 амінокислотних залишків), яка безпосередньо сполучається з рецепторною зоною антитіла.
Антитіло – це білкова молекула, яка специфічно взаємодіє з відповідним антигеном. Усі антитіла належать до гамма-глобулінів сироватки крові. Оскільки саме вони здійснюють специфічний імунітет, їх називають імуноглобулінами і за низкою ознак поділяють на 5 класів: IgM, IgG, IgA, IgE, IgD. Молекулярна маса імуноглобулінів становить 150-200 кД.
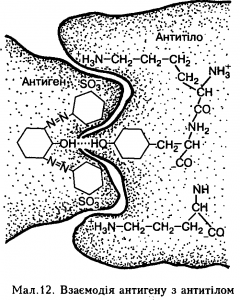
Структурноімуноглобулін (антитіло) складається з великої білкової молекули, в якій є відносно невелика (до 20 амінокислот) активна розпізнавальна група – рецептор, що забезпечує специфічність цього антитіла, яка ґрунтується на високій відповідності (комплементарності) структури активного центру антитіла та детермінантних груп антигену, причому відповідність має бути не тільки у просторовій конфігурації амінокислотних радикалів обох взаємодіючих молекул, а й у їхніх електричних зарядах (мал. 12).
Мал. 12