

ЗАНЯТИЕ № 3
ТЕМА: МОРФОЛОГИЯ СПИРОХЕТ, АКТИНОМИЦЕТОВ, ГРИБОВ, РИККЕТСИЙ, ХЛАМИДИЙ, МИКОПЛАЗМ, L-ФОРМ БАКТЕРИЙ
ПЕРЕЧЕНЬ КОНТРОЛЬНЫХ ВОПРОСОВ
-
Спирохеты. Систематическое положение и морфология спирохет. Особенности ультраструктуры и химического состава. Методы исследования.
-
Актиномицеты, морфология, ультраструктура, химический состав. Патогенные виды. Роль актиномицетов в природе и медицине. Методы выявления.
-
Таксономия хламидий. Морфология, структура, способы выявления. Цикл развития хламидий.
-
Риккетсии, морфология, ультраструктура, химический состав. Патогенные виды.
-
Микоплазмы. Классификация. Филогенез. Способы выявления.
-
Дефектные формы микробов: протопласты, сферопласты, L-формы.
САМОСТОЯТЕЛЬНАЯ И ЛАБОРАТОРНАЯ РАБОТА
1► Приготовление мазков из агаровой культуры дрожжеподобных грибов, окраска водным раствором фуксина, микроскопия.
2► Изучение морфологии плесневых грибов.
|
|
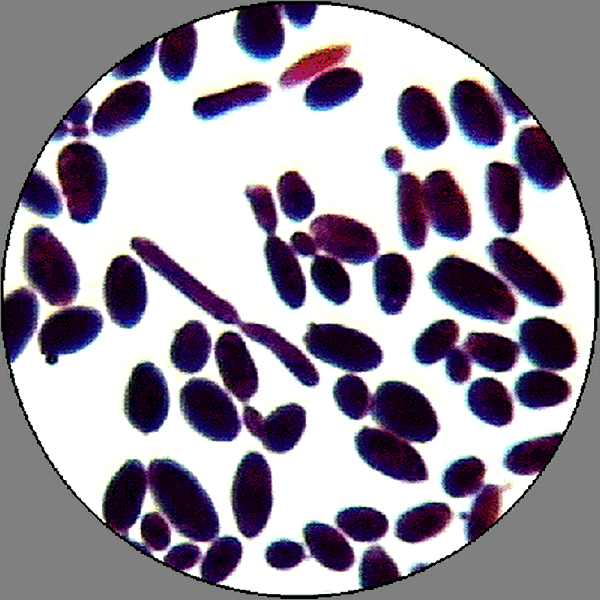
Candida albicans Окраска по Граму Увеличение х 630 Обозначьте на рисунке: 1- дрожжевая клетка; 2- псевдомицелий. |
|
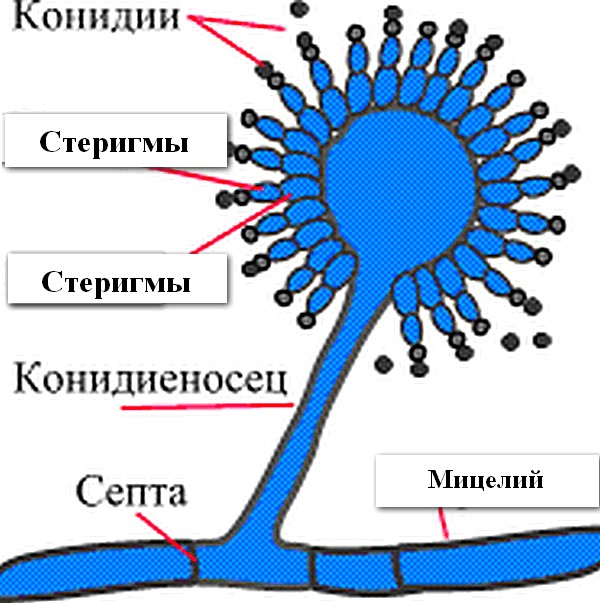
Препарат культуры плесневых грибов Окраска по по Граму Увеличение х 630 Обозначьте на рисунке: 1- конидии (спорангии); 2- стеригмы; 3- конидиеносец (спорангиеносец); 4- мицелий. |

3► Микроскопия и зарисовка риккетсий Провацека в готовых препаратах, окрашенных по Романовскому-Гимзе и Здродовскому.
ДЕМОНСТРАЦИИ
|
Rickettsia prowazekii в органах белых мышей |
Rickettsia prowazekii вакцинный штамм |
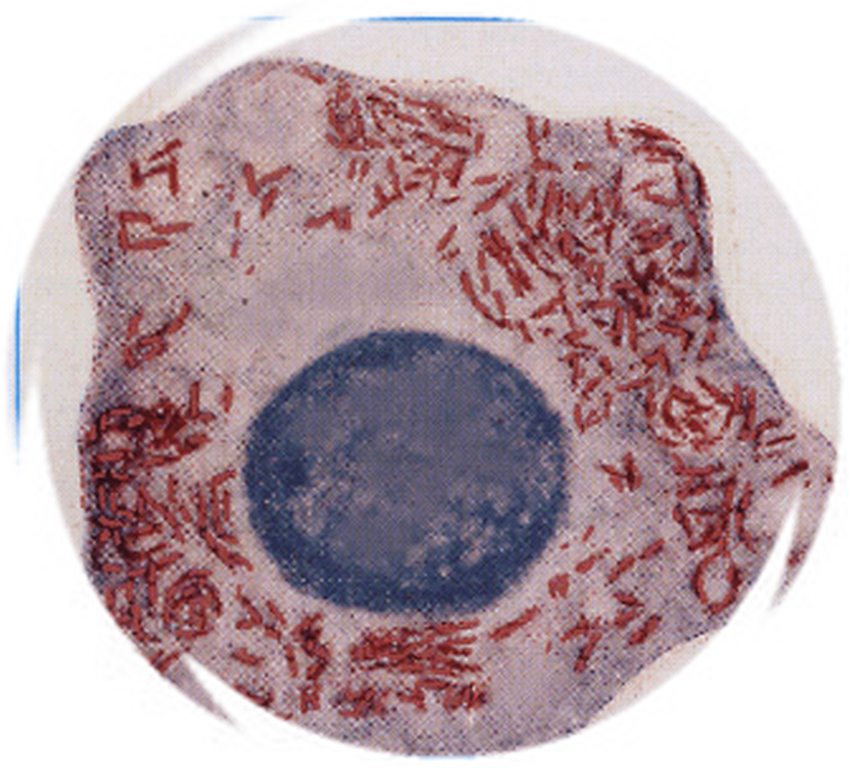
Borrelia recurrrentis в мазке крови |
|
|
|
|
|
Окраска по Здродовскому х 1000 |
Окраска по Здродовскому х 1000 |
Окраска по Романовскому-Гимзе х 1000 |
|
Палочковидные микроорганизмы, расположены внутриклеточно |
Вакцинный штамм представлен кокковидной формой риккетсий |
Боррелии - тонкие спиральные бактерии, диаметром 0,2-0,5 мкм, длиной 5,0-25 мкм, имеющие 3-10 крупных завитков.Окрашиваются по Романовскому - Гимзе в сине-фиолетовый цвет. |

(указать методы окраски микроскопических препаратов и особенности морфологии микроорганизмов – форму и расположение в препарате)
Тезисы
Определитель бактерий ("Bergey's Manual of Determinative Bacteriology") является справочником практических лабораторий и используется для идентификации бактерий по морфологическим, тинкториальным, биохимическим и антигенным свойствам.
Определитель бактерий Берджи делит царство прокариот на четыре отдела и 35 групп:
1.Gracilicutes - тонкостенные, грамотрицательные (1-16 группы);
2.Firmicutes - толстостенные, грамположительные (бактерии - 17-21 группы и актиномицеты - группы 22-29);
3.Tenericutes - лишены клеточной стенки (30-ая группа);
4.Mendosicutes - архебактерии - стенки лишены пептидогликана, имеются особенности строения рибосом, мембраны и РНК (31-35 группы).
Риккетсии грамотрицательные бактерии (от 0,3 до 0,5 мкм в диаметре и 0,8 до 2,0 микрометров в длину), облигатные паразиты клеток членистоногих и позвоночных. В организме человека риккетсии поражают эндотелий капилляров, что приводит к тяжелым заболеваниям, иногда с летальным исходом. Например, эпидемический возвратный тиф и его хроническая форма - болезнь Брилля-Цинссера (рецидивирующий возвратный тиф).
Грибы или Fungi. К этому царству относят 4 типа: Zygomycota (включают род Mucor, распространены в почве и воздухе, способны вызывать мукоромикоз легких, головного мозга и других органов человека и животных), Ascomycota (или сумчатые грибки, относятся к высшим грибам, к ним относятся роды Aspergillus, Penicillium, а также дрожжевые грибки), Basidiomycota (шляпочные грибы, а так же возбудители криптококкоза, разноцветного лишая, белой пьедры и трихоспороноза), Deuteromycota (несовершенные грибки, не размножаются половым путем, к ним относится род Candida). Внутри каждого типа, имеющего окончание -mycota, существуют основные таксономические деления: классы (-mycetes), порядки (-ales), семейства (-mycetaceae), роды и виды.
Филогенетически грибки близки к растениям, с которыми их сближает ряд общих признаков: наличие клеточной стенки и вакуолей, заполненных клеточным соком, хорошо видимое под микроскопом движение протоплазмы, характер поглощения питательных веществ, неспособность к активному перемещению, способность к неограниченному росту, необходимость прикрепления к субстрату, способ размножения и распространения спорами. С другой стороны, гетеротрофный тип питания, потребность в витаминах, способность к синтезу хитина, образование и накопление мочевины и гликогена (а не крахмала) придает им определенное сходство с животными клетками.
По строению грибки принято делить на дрожжи (Blastomycetes), плесени (Hyphomycetes) и шляпочные грибы. Возбудители микозов (заболеваний, вызванных грибками) относятся к первым двум группам. Среди грибков есть виды, которые в зависимости от условий растут либо как дрожжи, либо как плесени. Это явление называется диморфизмом, а такие грибки - диморфными. Диморфизм характерен для возбудителей системных микозов человека - бластомикоза (Blastomyces dermatidis, Paracoccidioides brasiliens), гистоплазмоза (Histoplasma capsularum) и кокцидиоидомикоза (Coccidioides immitis). В организме хозяина они образуют дрожжеподобные клетки, а в лабораторных условиях растут в виде мицелярных форм. Для Candida albicans характерно образование одновременно обеих форм. Большинство грибков способны как к половому, так и к бесполому способам размножения. Все структуры вегетативного и репродукционного бесполого размножения грибков называют анаморфами, тогда как структуры, образующиеся в результате полового процесса размножения, называют телеоморфами. Стадии телеоморфы и анаморфы характерны для всех грибков, кроме дейтеромицетов (несовершенных грибков), которым присуща только стадия анаморфы.
Заболевания, вызываемые патогенными грибками, называются микозами. Классифицируют микозы в зависимости от локализации процесса и агрессивности возбудителя против нормального или ослабленного организма. Выделяют поверхностные (кератомикозы), кожные (дерматомикозы), подкожные, глубокие (системные), оппортунистические микозы и микотоксикозы.
Протопласт (от греч. πρώτος — «первый» + греч. греч. πλαστός — образованный, вылепленный и др.) — содержимое растительной или бактериальной клетки, за исключением внешней клеточной оболочки (клеточной стенки), однако при сохранении клеточной (плазматической) мембраны. L-формы бактерий и микоплазмы изначально лишены клеточных стенок, по этому признаку их относят к протопластам.
Протопласт включает цитоплазму, ядро, все органеллы клеточную мембрану т.е протоплазму + мембрану.
Термин протопласт введен А. Ганштейном в 1880 году. Выделение протопластов широко используется в исследованиях клеток и генной инженерии. Жизнеспособные протопласты получают, в частности, в результате обработки клеток специальными ферментами, разрушающими клеточную оболочку. Это позволяет путем химических и механических методов выделять и исследовать структурные компоненты и даже отдельные молекулы, вносить изменения в генетическую структуру и так далее. Протопласты могут регенировать клеточную оболочку, что используется, в частности, для получения полноценных генетически измененных клеток.
После слияния (соматической гибридизации) протопласты образуют целые живые организмы — регенеранты. Таким путем могут быть получены соматические гибриды растений.
Клеточная стенка бактерий выполняет следующие функции: • механическую защиту клетки от воздействий факторов окружающей среды; • обеспечивает поддержание формы бактериальной клетки; • дает возможность клетке существовать в гипотонических растворах; • осуществляет транспорт веществ и ионов (характерно для грамотрицательных бактерий, имеющих наружную мембрану, которая является дополнительным барьером для их поступления; основным барьером служит цитоплазматическая мембрана); • препятствует проникновению в клетку токсических веществ (также более характерно для грамотрицательных бактерий, имеющих наружную мембрану); • на клеточной стенке находятся рецепторы, на которых адсорбируются бактериофаги и бактериоцины;
• в клеточной стенке находятся антигены (липополисахариды у грамотрицательных бактерий и тейховые кислоты у грамположительных бактерий); • на клеточной стенке находятся рецепторы, ответственные за взаимодействие клеток донора и реципиента при конъюгации бактерий.
Клеточная стенка не является жизненно важной структурой, так как она может быть удалена и бактериальные клетки при этом существуют в виде протопластов или сферопластов. Протопласты - клетки грамположительных бактерий, полностью лишенные клеточной стенки и окруженные только цитоплазматической мембраной.
Сферопласты образуются преимущественно из клеток грамотрицательных бактерий и отличаются от протопластов тем, что у них сохраняются остатки клеточной стенки.
Протопласты и сферопласты можно получить в лабораторных условиях, обрабатывая клетки бактерий лизоцимом, разрушающим муреин; антибиотиками пенициллинового ряда (пенициллин, ампициллин, карбенициллин и др.) или циклосерином, подавляющими синтез муреина. Фермент лизоцим действует на β-1,4-гликозидные связи муреина и тем самым разрушает его у бактерий со сформировавшейся клеточной стенкой. Антибиотики пенициллинового ряда и циклосерин оказывают действие только на растущие бактерии, нарушая синтез муреина клеточной стенки, именно они препятствуют поперечной сшивке пептидогликановых цепей, т. е. образованию пептидных связей. Протопласты и сферопласты можно получить и с помощью других ферментов, которые разрушают пептидные связи, участвующие в поперечной сшивке гетерополимерных цепей муреина. В качестве примера можно привести фермент эндопептидазу, синтезируемую бактериями Escherichia coli. Этот фермент разрывает пептидную связь между D-аланином и мезо-диаминопимелиновой кислотой. Протопласты и сферопласты стабильно сохраняются в гипертонических или изотонических условиях. Для создания гипертонических условий чаще всего используют сахарозу или маннит в концентрациях 0,1–1,0 М. В гипотонических условиях протопласты и сферопласты лопаются и образуют «тени».
Протопласты и сферопласты в 3–10 раз крупнее исходных клеток бактерий. В гипертонических или изотонических условиях они осуществляют обмен веществ, характерный для исходных клеток, т. е. сохраняют дыхательную активность, синтезируют необходимые биополимеры, образуют эндоспоры, если процесс споруляции уже был инициирован. Можно наблюдать рост сферопластов и протопластов, а иногда и их деление. В отличие от исходных клеток, на них не адсорбируются бактериофаги и бактериоцины. Кроме того, у протопластов и сферопластов отсутствуют мезосомы – производные цитоплазматической мембраны. При снятии действующего на образование муреина фактора (пенициллин, циклосерин, лизоцим и др.) протопласты, как правило, отмирают, реже регенерируют клеточную стенку и возвращаются в исходное состояние, но могут превращаться в L-формы. Сферопласты ревертируют (превращаются) в нормальные бактериальные клетки, либо превращаются в L-формы, либо отмирают.
Бактерии, частично или полностью лишенные клеточной стенки, но сохранившие способность к развитию, принято называть L-формами. Буква L – первая буква названия Листеровского института в Лондоне, где впервые обратили внимание на развитие морфологически весьма необычных клеток в культуре бактерий Streptococcus moniliformis, выделенной из жидкости уха крысы. Позже были описаны L-формы у самых разных видов бактерий. Было показано, что L-формы возникают спонтанно или индуцированно – под воздействием агентов, блокирующих синтез клеточной стенки (антибиотиков пенициллинового ряда и циклосерина, ультрафиолетовых и рентгеновских лучей, аминокислоты глицина). L-формы образуются в результате несбалансированного роста нормальных бактериальных клеток в длину и толщину и поэтому плеоморфные. В культурах L-форм обнаруживаются клетки размером 0,2–50 мкм. Они шаровидные, нитевидные, присутствуют и бесструктурные массы. L-формы проходят через бактериальные фильтры и легко разрушаются при механических воздействиях. В отличие от протопластов и сферопластов, клетки L-форм имеют хорошо развитую систему внутрицитоплазматических мембран, т. е. у них содержатся мезосомы, а в отличие от нормальных клеток L-формы часто содержат крупные вакуоли. L-формы обладают пониженным уровнем метаболической активности по сравнению с исходными бактериями. Они нечувствительны к любым агентам, действующим на клеточную стенку. Культивировать L-формы можно только на специальных средах с высоким осмотическим давлением. L-формы лучше растут на плотной, чем в жидкой среде. На плотной среде они образуют колонии, врастающие в агар и имеющие характерную форму перевернутой шляпы. Колонии растут медленно, хотя иногда достигают значительных размеров. У L-форм не функционируют нормальные механизмы клеточного деления. В основном они делятся с образованием элементарных тел, которые отпочковываются от поверхности клетки или от мембраны вакуоли. Различают стабильные и нестабильные L-формы. Нестабильные L-формы обладают элементами клеточной стенки и поэтому способны ревертировать в нормальные бактериальные клетки после исключения действия фактора, вызвавшего их образование. Стабильные L-формы полностью лишены ригидной клеточной стенки, что сближает их с протопластами. Они крайне редко ревертируют в исходные бактериальные формы и существуют без изменений в различных условиях среды. Переход в L-форму можно рассматривать как способ переживания бактериями неблагоприятных условий, особенно у патогенных микроорганизмов. Исследования L-форм представляют существенный интерес для медицинской микробиологии, поскольку в таком состоянии в организме человека и животных могут сохраняться патогенные бактерии. При нерациональном использовании антибиотиков, приводящем к образованию L-форм из бактерий, может наступить кажущееся улучшение состояния больного. Однако после прекращения приема лечебного препарата происходит превращение L-форм в бактерии исходного вида с восстановлением их вирулентности, что приводит к рецидиву болезни.
Бактерии, у которых отсутствует клеточная стенка, существуют и в природе: это микоплазмы. Первым описанным представителем микоплазм явился возбудитель плевропневмонии крупного рогатого скота. Подобные микроорганизмы обнаружены и у других животных – овец, коз, крыс, собак, а также у человека, всем им было дано общее название РРLО (плевропневмониеподобные организмы). Кроме того, микоплазмы могут существовать как сапрофиты в естественных условиях, а также вызывать заболевания и у растений.