

Особенности сна у животных
Стадия быстрых движений глаз и выраженный рисунок сна есть только у теплокровных — млекопитающих (кроме ехидны) и птиц, а также высших пресмыкающихся (Латаш, 1975). Полагают, что картина сна в эволюции формировалась раздельно у млекопитающих и птиц; у последних длительность парадоксального сна меньше, а число циклов больше (Моисеева, 1981). Чем короче циклы сна и более длителен сам сон, тем короче жизнь животного (Борбели, 1989). Особенностью цикличности сна животных является то, что они просыпаются по окончании каждого цикла сна, что позволяет им быть бдительными по отношению к хищникам.
Внешнее проявление быстрого сна у животных несколько отличается от того, что наблюдается у человека. Появление сновидений сопровождается подергиванием лап и ушей, движением хвоста и челюстей, повизгиванием.
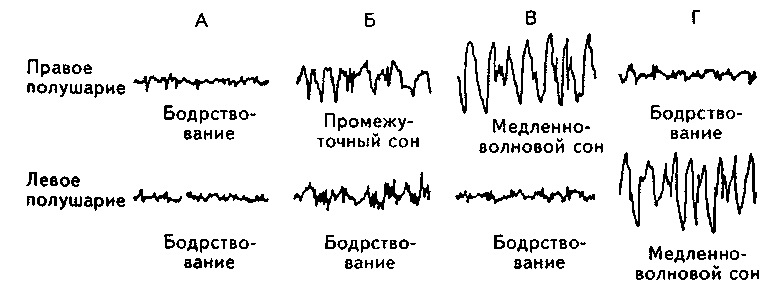
У морских млекопитающих (например, дельфинов), которые должны выныривать, чтобы сделать вдох, отмечен либо однополушарный сон, когда в одном полушарии могут наблюдаться медленные волны, а в другом — ЭЭГ бодрствования, либо все стадии сна укладываются в период между двумя вдохами (например, у тюленей) (рис. 9.5).
Наблюдения за животными различного размера и веса привели к возникновению гипотезы, что сон у млекопитающих укорачивается при нарастании массы тела (Elgar e. а., 1988; Zepelin, 1994). Однако, используя современные методы регистрации свободного поведения животных (такие как ви-деофильмирование в течение суток), исследователи выявляют значительные периоды дневного сна у крупных травоядных, что ставит под сомнение эту гипотезу (Tobler, Schwierin, 1996).
Рис. 9.5. Сон ДМъфина (Mukhametov, 1984).
228
Если сон в течение суток несколько раз сменяется бодрствованием, то он называется полифазным. Такой тип сна отмечается у маленьких детей и некоторых
Для животных характерен еще один вариант сна — спячка. Животные, проживающие в условиях, когда они не могут обеспечить себя кормом в определенный период (на севере из-за снега, в пустынях — из-за усиленной жары и отсутствия воды), засыпают на все время отсутствия еды. При этом у них изменяется теплообмен за счет некоторого понижения температуры тела, замедления обмена веществ, дыхания и сердцебиения.
Гипотезы, объясняющие причины сна
Существует несколько гипотез о причинах возникновения сна и сновидений у человека. Наиболее старая объясняет возникновение сна накоплением неких “сомногенных” факторов в течение времени бодрствования. Неоднократно проводились исследования, в которых бодрствующим животным давали экстракты из крови или спинномозговой жидкости долго не спящих животных. Получив такой препарат, последние становились сонливыми и утомленными. В то же время наблюдения над сиамскими близнецами, имеющими единую систему кровообращения, не соответствовали этим данным. П.К. Анохин отмечал, что одна голова близнецов могла бодрствовать, когда другая спала.
Многие вещества проверялись в качестве таких факторов, однако подтвердить гипотезу долго не удавалось. Согласно нейромедиаторной гипотезе, причина сна связана с ритмическими изменениями содержания медиаторов (Jouvet, 1988). Современный вариант ее основан на изменении концентрации аденозина в течение дня. Х.Х. Бенингтон и Х.С. Хеллер (Benington, Heller, 1995) предложили биохимическую модель сонного гомеостаза: во время сна пополняются запасы мозгового гликогена, который интенсивно истощается в астроцитах в течение дня. Отсутствие гликогена инициирует выброс во внеклеточное пространство аденозина, который действует как нейромодулятор, обеспечивающий начало сна через появление медленных волн. Он усиливает обратный ток калия к нейронам, что снижает их мембранный потенциал, вследствие чего формируются медленные волны.
Циркадная гипотеза опирается на идею о внешнем контроле со стороны вариаций средовых факторов (Bixler, Vela-Bueno, 1987).
Сон есть активный феномен, являющийся следствием взаимодействия особых нейронов, генерирующих сон, с мозговыми центрами жизненно важных функций. Цикл сна выработался в эволюции как механизм, облегчающий реакцию на опасность. Генератор быстрого сна — это группа нейронов ствола и базальных ядер переднего мозга.
Согласно представлению Н. Моисеевой (1985), тот факт, что быстрый сон независимо появился в двух ветвях теплокровных животных — у птиц и млекопитающих, свидетельствует о его эволюционной значимости. На основании данных, указывающих на некоторую функциональную разобщенность структур мозга и его элементов, ученая предположила, что в процессе сна преиму-
229

Нейрофизиологическая гипотеза приписывает возникновение быстрого сна активному механизму, имеющему периодическое нарастание и спад деятельности. По-видимому, в это время включаются в работу нейроны, незадействованные в течение дня. Известно, что гибель нейронов напрямую зависит от их функциональной активности. Таким образом, сон может способствовать сохранности нейронов мозга (Годфруа, 1992). И.Н. Пигарев (Pigarev, 1994) обнаружил, что во сне ряд структур меняет тип своей активности. Например, те области мозга, которые обрабатывают экстероцептив-ную (идущую извне) информацию днем, ночью переключаются на интеро-цептивную информацию. Возможно, это и есть отражение механизма, сохраняющего существование временно не работающих нейронов.
До сих пор неясна функция сновидений: возможно, это механизм разрядки напряжения, идущего из бессознательного, или способ освобождения от избытка информации для упрочения следов наиболее значимой; наконец, они могут быть проявлением случайной активности нейронов. Особая форма сновидений — люцидные сновидения, при которых человек осознает, что он спит, не находит объяснения.
Сновидения, согласно психоаналитической концепции, принимают участие в формировании психологической защиты. Стрессовые стимулы при этом включаются в содержание сновидения в форме, снижающей их травмирующее значение (Латаш, 1975). Есть гипотеза, что в это время происходит вытеснение информации, которая не совместима с жизненным опытом, человека и его морально-этическими установками (Моисеева, 1981).
Существует гипотеза анаболической функции сна, согласно которой в третьей и четвертой стадиях сна идут процессы синтеза в организме в целом, а парадоксальный сон представляет собой состояние, обеспечивающее синтез в мозге. Доказательством этой гипотезы могут быть факты выделения гормона роста в дельта-сне (Takahashi, 1979). Однако есть и достаточное количество возражений, поскольку для реализации действия гормона роста необходимы свободные аминокислоты, которые могут быть в крови только в течение первых пяти часов после еды. Известно, что большинство людей ест за несколько часов до сна, следовательно, гормон роста не может оказать свое физиологическое действие (Ноте, 1988). В любом случае остается еще много темных пятен в вопросах о причинах столь обыденного явления, как сон.