

Цикл сон — бодрствование
Обращает на себя внимание ритмичность многих процессов в организме. Цикл сон — бодрствование постоянно подстраивается под внешние датчики (длительность дня и ночи) и составляет 24 часа; синхронно ему настроены другие ритмы тела — гормональный, физиологический, эмоциональный, поведенческий. Биологические часы расположены, по-видимому, в гипоталамусе (в супрахиазматическом ядре) и эпифизе (Zucker, 1980).
Биологический цикл несколько больше 24 часов, поскольку в тех случаях, когда человек находится в изолированном помещении и не имеет возможности подстраиваться под внешние часы (восход и заход солнца), его внутренний цикл сон — бодрствование составляет около 30 часов (Aschoff, 1965; Putilov, 1995). Впервые это показал французский спелеолог Мишель Сифр (Siffre, 1963), который провел в глубокой пещере под землей около 60 дней. Все это время он не имел никакой связи с наземным миром, хотя специальные датчики, поставленные на нем, передавали информацию о его цикле сон — бодрствование. Через какое-то время цикл стабилизировался и составил более 25 часов.
В естественных условиях биологические часы регулярно синхронизируются с циклом день — ночь. Соответствие световой информации биологическим часам контролируется эпифизом. Информация об интенсивности света от ретины передается, с одной стороны, в зрительную кору, с другой — в гипоталамус и эпифиз. Эпифиз ритмично вырабатывает гормон мелатонин; этот процесс в настоящее время рассматривается как аналог биологических часов (Putilov, 1995).
Уровни активации и эффективность психических процессов
P.M. Йеркс и Дж.Д. Додсон (Yerkes, Dodson, 1908) в начале века открыли закон, согласно которому для каждого типа заданий есть свой оптимум активации (определенное благоприятное состояние напряжения), обеспечивающий наиболее успешное его исполнение. Графически этот закон можно описать колоколообразной кривой, которую также называют обратной U-образной зависимостью. На практике она проявляется в низкой эффективности любой деятельности (обучения, внимания, операторской работы, мышечной активности и т. д.) как при возбуждении, так и при полном расслаб-
203
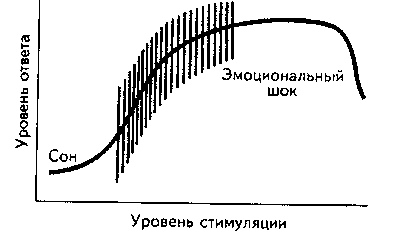
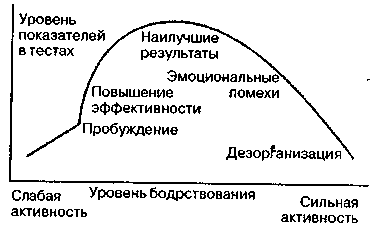
Рис. 8.3. Зависимость эффективности деятельности от уровня активации (Фресс, 1975).
На эти данные обратили внимание лишь после более поздней работы Е. Даффи (Duffy, 1932), которая установила, что эффективность выполнения психологических тестов у детей ниже в случае высокого уровня мышечного напряжения (измеряемого посредством динамометра) по сравнению со средними значениями этого показателя.
Рис. 8.4. Зависимость эффективности поведения от уровня активации. Видоизмененный Д.О. Хеббом закон Йеркса-Додсона (Годф-руа, 1992).
В дальнейшем данная зависимость неоднократно подтверждалась (Иваницкий, 1976; Зыков, Мелехова, 1976). В экспериментах с записью психофизиологических коррелятов активационных процессов было показано, что слова, вызывающие у испытуемого большую амплитуду КГР или большую частоту сердечных сокращений (ЧСС) (воспринятые на более высоком уровне активации),
лучше сохраняются в памяти (O'Gorman, Jameson, 1978; Plouffe, Stelmack, 1982, 1984; Conte, Kinsbourne, 1988).
В то же время ряд исследований не подтверждает существование этой простой закономерности (Santley, LaCava, 1977). Подобное расхождение в результатах можно объяснить тем, что зависимость запоминания от уровня активации подвергается действию множества других влияний (мотивация, личностные особенности и т. д.), и конечный результат исследования зависит от общей специфики построения эксперимента.
Было высказано предположение, что между уровнем активации и шкалированием эмоций также существует зависимость, подчиняющаяся закону Йеркса-Додсона (Breitkopf, 1982). На практике это означает, что максимальная вариабельность эмоциональных ответов испытуемых должна наблюдаться при оптимуме активации. Д.О. Хебб (1955) проиллюстрировал это графически (рис. 8.4).
Исследования, проведенные в школе, показали, что успевающие ученики работают в оптимальном уровне активации, поэтому путем изменения функционального состояния ребенка (контролируя это, например, по величине КГР) можно улучшить обучение отстающих детей (Mangina, 1989).