

Сетчатка и ее функции
На сетчатой оболочке глаза имеется два типа рецепторов — палочки (около 120 млн) и колбочки (6 млн). Каждый из них состоит из внутреннего членика, напоминающего обычную нервную клетку, и палочко- или кол-бочкообразного наружного членика, чувствительного к свету (рис. 5.11).
Внутренний членик необходим для поддержания жизнедеятельности клетки, наружный реагирует на свет благодаря находящемуся в нем пигменту. Наружный членик состоит из множества дисков, лежащих стопками, один на другом. Одна палочка содержит до миллиона таких дисков. Свет сначала проходит сквозь внутренний членик, а затем попадает в наружный, где он захватывается фоточувствительными молекулами, расположенными на дисках.
Колбочки являются рецепторами цветового зрения и возбуждаются при ярком свете. Палочки активируются в сумерках и способствуют возникновению ощущения серого цве-
та, именно поэтому ночью все предметы воспринимаются как серые.
Каждая колбочка связана с мозгом отдельным волокном, и они функционируют по отдельности. Палочки работают группами, от каждой из которых
128
129
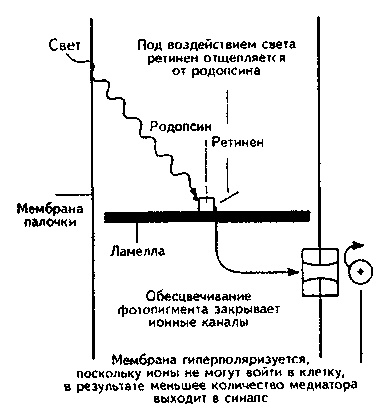
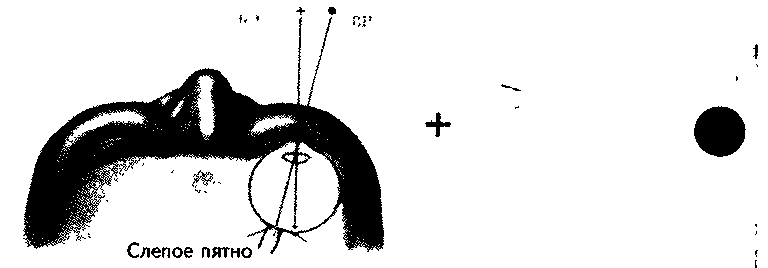
Рис. 5.12. Тест на слепое пятно. Закройте левый глаз и смотрите на знак “+” правым глазом. Приближайте страницу к глазу до тех пор, пока круг не исчезнет, поскольку его изображе* ние попадает в слепое пятно (Carlson, 1992).
только одно волокно входит в состав зрительного нерва. Палочки активируются светом умеренной интенсивности, что сопровождается появлением слабого ощущения цвета.
Рецепторы распределены по сетчатке неравномерно. В области центральной ямки находятся в основном колбочки (до 140 тыс. на 1 мм2 поверхности). По направлению к периферии число колбочек уменьшается, а число палочек растет. Место входа зрительного нерва — сосок зрительного нерва — совсем не содержит рецепторов и нечувствительно к свету, поэтому называется слепым пятном. Обычно человек не замечает слепого пятна, но его наличие можно продемонстрировать экспериментом (рис. 5.12).
Фотопигменты — опсины, — найденные в органах зрения человека и животных, имеют в своей основе витамин А, связанный с белками. В палочках выявлен пигмент родопсин, состоящий из опсина и альдегида витамина А — ретинена. Он обеспечивает максимум поглощения световых лучей с длиной волны 500 мкм, что относится к зеленой части спектра. Именно зеленые предметы кажутся наиболее яркими в темноте.
Существуют три типа колбочек, содержащих различные пигменты, чувствительные к синему, зеленому и красному свету. Например, пигмент, реагирующий на красный цвет, называется йодопсином. В пользу того, что нормальное трихроматическое цветовое зрение основано на трех независимых друг от друга рецепторных механизмах, говорит существование трех видов цветовой слепоты, проявляющихся при отсутствии одного из рецепторов.
Если обработать сетчатку раствором квасцов, родопсин не будет более распадаться, и на сетчатке можно увидеть изображение предмета, на который смотрел глаз. Полученные таким образом снимки В. Кюне называл оптографией. Он впервые в 1880 г. исследовал оптограмму казненного преступника (рис. 5.13). В. Кюне приложил много труда, чтобы определить, что изображает эта опто-грамма, но его усилия не увенчались успехом (Уолд, 1974). Однако до сих пор идея опознания преступника по изображению на сетчатке жертвы в момент совершения преступления мелькает на страницах детективных историй.
При поглощении кванта света зрительным пигментом его боковая цепь выл-
130
рямляется, и он превращается в свои изомер, в результате чего нарушается его связь с белком. Мембраны фоторецепторов отличаются от мембран других нейронов, поскольку их натрий-калиевые каналы постоянно открыты. Это приводит к тому, что некоторое количество медиатора постоянно выходит в синаптическую щель. Следовательно, эта мембрана менее поляризована, чем окружающие. При расщеплении пигмента каналы закрываются, мембрана гиперполяризуется и выход медиатора прекращается (рис. 5.14) (Carlson, 1992).
Рис. 5.13. Оптограмма человека на рисунке Кюне; так и не удалось определить, что она изображает (Уолд, 1974).
Рецепторная клетка соединена с биполярной. Фоторецепторы и биполярные клетки сами не создают потенциала действия. Их обмен медиатором регулируется мембранным потенциалом. Деполяризация увеличивает его количество, гиперполяризация— уменьшает (рис. 5.15). Поскольку в норме медиатор гиперпо-ляризует мембрану дендритов биполярных клеток, уменьшение медиатора ведет к ее деполяризации. Таким образом, свет гиперполяризует рецепторную мембрану и вызывает деполяризацию мембраны биполярных клеток. Эта деполяризация ведет к выбросу медиатора в синапсе между биполярной и ганглиозной клетками, вызывая изменение ее импуль-сации и, следовательно, изменение сигнала, поступающего в мозг.
Рис.
5.14. Процесс
превращения света в электрический
сигнал в фоторецепторах (Carlson,
1992).
131
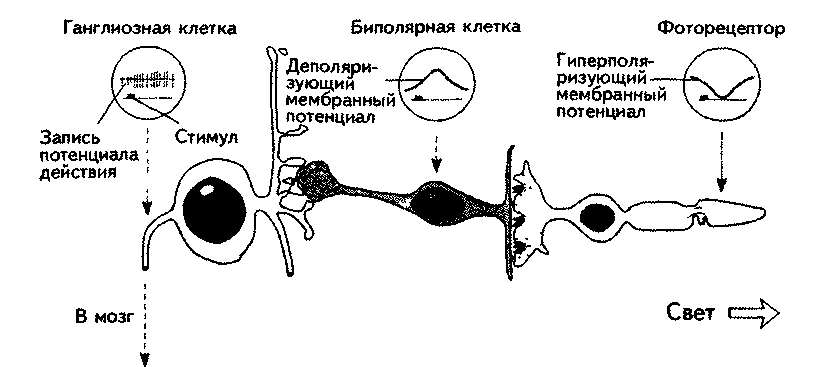
сетчатки, с которого информация поступает в одну ганглиозную клетку, называется рецептивным полем этой клетки (Хьюбель, 1974).
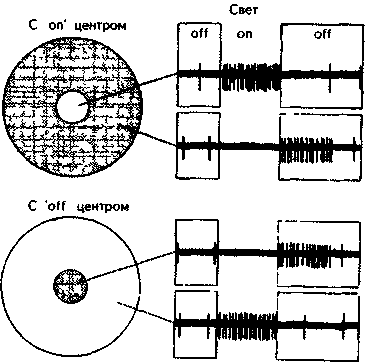
Рис. 5 16. Концентрические рецептивные поля характерны для ганглиозных клеток сетчатки и клеток коленчатого тела На верхней осциллограмме показана сильная импульсация клетки с “оп”-центром при освещении центра поля, если свет падает на “off”-зону, то импульсация прекращается до тех пор, пока свет не выключат Внизу показана реакция клетки с “оп~”-центром (Хьюбель, 1974).
Ганглиозные клетки сетчатки разряжаются в постоянном ритме даже при отсутствии всякого раздражения. Если частота импульсации ганглиозной клетки под воздействием света нарастает, то реакция называется “оп”-ответом, если импульса-ция падает, то говорят об “оп"”-отве-те. Отделы сетчатки, с которых вызываются эти реакции, соответственно называются “on”- и “off”-участками. В свою очередь, гангли-озные клетки, получающие информацию от этих клеток, делятся на клетки с “on”- и “off”-центрами (рис. 5.16). Клетки с “оп”-центрами имеют небольшой “on”-участок и окружающий его “offV-участок. Напротив, клетки с “offr-центрами
имеют центральный “offV-участок и “оп”-периферию (Kuffler, 1953).
Рецептивные поля ганглиозных клеток производят поточечное описание изображения на сетчатке. Если несколько рядом расположенных ганглиозных клеток активируются вместе, это приводит к их взаимному торможению, которое называется латеральным (боковым).