

21 Глия
Нейроны составляют лишь 25% от всех клеток мозга, остальные 75% клеток относятся к нейроглии (glia — клей, греч.). Это название было дано в 1846 г. Р. Вирховым, полагавшим, что глия - это цементирующая основа для объединения нервных клеток. В среднем глиальные клетки составляют по величине примерно 1/10 размера нейрона. В отличие от нейронов они способны делиться. Именно благодаря им происходит увеличение объема мозга ребенка, составляющего при рождении примерно четверть мозга взрослого. Возникновение опухолей в мозге также связано не с активностью нейронов, а с бесконтрольным делением глиальных клеток.
Глиальные клетки имеют множество функций, но они не передают информацию, как это делают нейроны (рис. 1.14). Мембранный потенциал глиальных клеток выше, чем у нейронов, и определяется разностью концентраций ионов калия во внутри- и внеклеточном пространстве. Это отличает их от нейронов, мембранный потенциал которых формируется как разностью концентраций ионов калия, так и ионов натрия. При возбуждении нейрона из него одновременно выходят ионы К+и Na+, что ведет к изменению мембранного потенциала расположенных рядом глиальных клеток. Последние частично поглощают ионы калия, функционируя как калиевый буфер, поддерживающий постоянную внеклеточную концентрацию этих ионов. Повышение внеклеточной концентрации калия могло бы снизить порог возбуждения нейрона, что вело бы к его спонтанной активации. Возможно, что именно этот механизм включается при возникновении эпилептических припадков (Хухо, 1990). Выделяют следующие функции глии.
Рис. 1.14. Формы некоторых глиальных клеток.
Два типа глиальных клеток образуют миелиновую оболочку для аксо нов: олигодендроциты формируют ее в головном и спинном мозге, а Шван- новские клетки — в периферической нервной системе. Они обертываются вокруг аксона, изолируя его и ускоряя проведение импульса. Отростки од ной глиальной клетки обертываются вокруг разных аксонов, что может спо собствовать интеграции работы сразу нескольких нейронов.
Астроглия и микроглия очищают мозг от погибших нейронов и от не нужного материала.
А
 строглия
также имеет опорную
функцию, заполняя промежутки
между
нейронами.
строглия
также имеет опорную
функцию, заполняя промежутки
между
нейронами.Радиальная глия помогает миг рации нейронов и направляет аксо ны в сторону расположения их ми шеней в период эмбрионального развития. Аналогичным образом Шванновские клетки при поврежде ниях направляют восстанавливаю щийся аксон к месту иннервации. Они участвуют и в востановлении поврежденных нервов. Было показа но, что после повреждения аксона Шванновская клетка может заменять утраченное нервное окончание в мышце и даже выделять медиатор (Хухо, 1990). В зрелом мозге ради альная глия перерождается в другие виды глии, осуществляя опорную функцию.
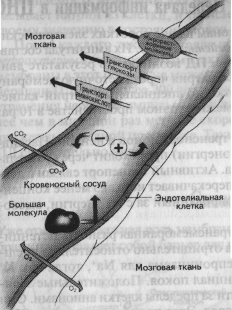
Рис. 1.15. Гематоэнцефалический барьер (Kalat, 1992).
5. Астроглия формирует уникальный защитный слой между нейроном и кровеносным сосудом, так что все вещества из крови могут попасть в нейрон только через глиальную клетку. Этот барьер называется гематоэнцефалическим (haima — кровь, enkephalos — мозг, греч.}. Гематоэнцефалический барьер могут преодолевать только маленькие молекулы, например ионы, глюкоза, незаменимые аминокислоты и жирные кислоты (рис. 1.15). Благодаря этому большие молекулы, токсины, вирусы и микробы не могут проникнуть в нейрон, что приводит к значительному повышению толерантности (устойчивости) мозга к вирусным инфекциям.
Существует только одна область мозга, где происходит нарушение гемато-энцефалического барьера, — гипоталамус. В нем находятся клетки, секрети-рующие либерины и статины, управляющие выделением гормонов из гипофиза. Сосуды непосредственно подходят к секретирующим нейронам, выделяющим свои биологически активные вещества прямо в кровь. Ввиду функциональной необходимости Гематоэнцефалический барьер в этом месте нарушается. Гипоталамус можно назвать “ахиллесовой пятой” мозга, поскольку только здесь возможно проникновение инфекций в нервную систему человека.
Наличие гематоэнцефалического барьера при инфекционных поражениях мозговой ткани может препятствовать ее лечению путем введения антибиотиков в кровь. Молекулы лекарства не могут попасть в мозг в нужном количестве и не имеют возможности подойти непосредственно к очагу инфекции. Единственным выходом из этой ситуации остается пункция: лекарство вводится в позвоночный канал, связанный с желудочками мозга, через которые и попадает к очагу инфекции.
22
23