

204 Роль ретикулярной формации среднего мозга в формировании состояния бодрствования
Физиологической основой градаций функциональных состояний является активность восходящей ретикулярной формации среднего мозга. Ее роль в регуляции цикла сон — бодрствование была продемонстрирована в 1938 г. Ф. Бре-мером, который показал, что у кошки с перерезанным мозговым стволом на уровне несколько ниже продолговатого мозга или варолиева моста (препарат encephale isole) сохранялась электрическая активность бодрствования. Перерезка мозга на уровне среднего мозга (препарат cerveau isole) вызывала коматозное состояние с регистрацией медленных высоковольтных колебаний (рис. 8.5). В 1949 г. Г. Моруцци и Г. Мэгун (Moruzzi, Magoun, 1949) доказали, что возникновение коматозного состояния обусловлено разъединением мезенцефалической ретикулярной формации и коры мозга. Высокочастотная электрическая стимуляция мезенцефалической ретикулярной формации вызвала реакцию активации на ЭЭГ. Низкочастотная стимуляция, напротив, давала противоположный эффект.
Анатомическая особенность ретикулярной формации заключается в том, что все восходящие сенсорные пути оставляют в ней свои коллатерали (боковые ветви). Это предопределяет возможность ее активации под воздействием любого стимула (рис. 8.6). Активированная ретикулярная формация, в свою очередь, возбуждает кору больших полушарий непосредственно и косвенно через другие структуры, в том числе ядра таламуса (Morgane, Stern, 1974).
Запись электрической активности нейронов ретикулярной формации с помощью микроэлектродов подтвердила причинную связь между возбуждением ее нейронов и активацией коры головного мозга.
Ретикулярная формация представляет собой сложную структуру из нейронов, имеющих длинные дендриты с многочисленными синаптическими

Рис. 8.5. Локализация систем, играющих критическую роль в регуляции цикла сон — бодрствование в мозге кошки. Сечение на уровне А ведет к постоянному сну у кошки. Сечение на уровне Б ведет к пролонгированному бодрствованию (Kalat, 1992).
205
связями (reticulum — сеть, лат.). Запись активности единичных нейронов мезенцефалической ретикулярной формации и ее сопоставление с изменениями на ЭЭГ у свободно двигающихся кошек с вживленными микроэлектродами (Siegel, 1979) не подтвердили гипотезы, что ретикулярная формация инициирует реакцию активации. Более того, установлено, что возбуждение отдельных нейронов тесно связано с движением глаз, ушей, мышц лица, тела, конечностей. Не обнаружилась корреляция индивидуальной активности с общим функциональным состоянием и циклом сон — бодрствование. Дж.М. Сигел предположил, что это несоответствие результатам других исследователей может иметь две причины (Siegel, 1979). Его предшественники использовали анестезированных животных, находящихся под действием наркотика в специальных станках. Невозможно наблюдать связь между движением и активностью единичных нейронов у обездвиженных животных. В тех же случаях, когда в эксперименте принимали участие движущиеся животные, делалась оценка записи электрической активности многих нейронов одновременно, что давало картину усреднения ПД. Возбуждение одного нейрона обусловлено конкретной реакцией животного, тогда как ответ группы нейронов является неспецифичным.
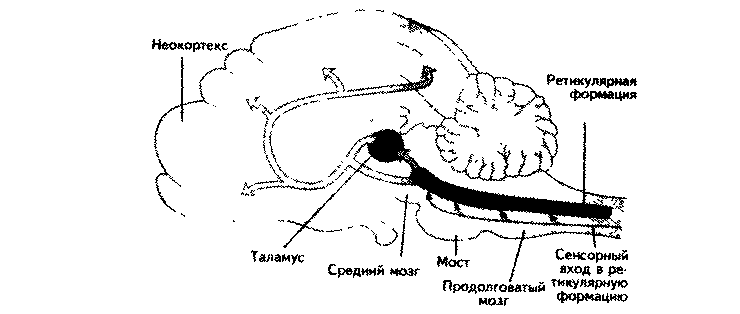
Современная точка зрения на связь активности мезенцефалической ретикулярной формации с реакцией активации в коре больших полушарий содержит несколько более ранних гипотез. Ретикулярная формация первично включена в контроль специфических действий, но отдельные ее ядра имеют непосредственный активирующий эффект на кору мозга. Например, введение агониста катехоламинов — амфетамина — вызывает реакцию активации в коре и бессонницу. Этот эффект опосредован норадренергичес-кой системой голубого пятна (locus coeruleus), расположенного в дорсальной части моста и являющегося частью ретикулярной формации (рис. 8.7). Нейроны этого ядра посылают широко ветвящиеся аксоны в неокортекс, гиппокамп, таламус, кору мозжечка, мост, продолговатый мозг, что подтверждает его способность воздействовать на эти значимые структуры мозга (Steriade, 1996).
206

При сопоставлении записи электрической активности нейронов голубого пятна и поведенческой активности у свободно передвигающихся крыс была обнаружена четкая корреляция между ними (рис. 8.8). Установлено, что пробуждение животного природными стимулами во время сна или повышение активности в период спокойного бодрствования ведет к возбуждению нейронов голубого пятна. Можно допустить, что нейроны этой структуры
Рис. 8.7. Ствол мозга человека (Carlson, 1992).
вызывают реакцию активации в коре. Однако их активность у свободно передвигающихся крыс с вживленными в эту область электродами крайне низка в процессе умывания или питья подслащенной воды — т. е. в поведении, которое соответствует высокому уровню бодрствования животного. Дж. Астон-Джонс и Ф.Е. Блум (Aston-Jones, Bloom, 1981) сняли это противоречие, предположив, что активность нейронов голубого пятна коррелирует скорее с бдительностью, т. е. ответом на внешние воздействия, чем с неспецифической активацией. Когда крыса реагировала на внешние
Средняя частота активности нейронов

Рис. 8.8. Активность норадренергических нейронов в locus coeruleus у свободно передвигающейся кошки в различные стадии цикла сон — бодрствование (Aston-Jones, Bloom, 1981).
207
стимулы, активность нейронов голубого пятна была высока, но во время умывания и питья животное в меньшей мере обращало внимание на окружение, что и приводило к снижению импульсаиии.