

Анализ зрительной информации
Впервые вопрос о специфике зрительного восприятия был сформулирован в рамках гештальт-психологии. Было отмечено, что обнаружение предмета начинается еще до того, как происходит опознание объекта. Современные представления о структурах мозга, анализирующих зрительную информацию, сформировались практически затри последних десятилетия.
В конце XIX века немецкий психиатр П. Флексиг из Лейпцигского университета показал, что определенные области головного мозга, в том числе стриарная кора, уже при рождении выглядят зрелыми, тогда как другие (ассоциативная, или престриарная кора) продолжают развиваться, как если бы их созревание зависело от приобретаемого опыта.
Функциональная организация коры головного мозга связана с существованием модулей — колонок, которые представляют собой вертикально расположенные группы клеток с многочисленными связями между ними в вертикальном направлении и незначительным числом связей в горизонтальном направлении (Мауткасл, 1981).
В своих физиологических исследованиях С. Зеки (1992) предъявлял макакам набор стимулов (цвета, линии различной ориентации и точки, движущиеся в разных направлениях) и с помощью электродов регистрировал активность нейронов престриарной коры. Престриарная зрительная кора включает несколько зон, отделенных от стриарной коры (зона VI) зоной V2. Оказалось, что в зоне V5 или МТ (рис. 5.19), все нейроны ответственны за восприятие движения, причем в большинстве случаев строго определенного направления, и никогда не реагируют на цвет движущегося стимула. Это навело автора на мысль о специализации зоны V5 на видении движения. В то же время подавляющее большинство клеток зоны V4 достаточно избирательно реагируют на свет той или иной длины волны, а многие из них специализированы и на определенной ориентации линий. Почти все клетки
135
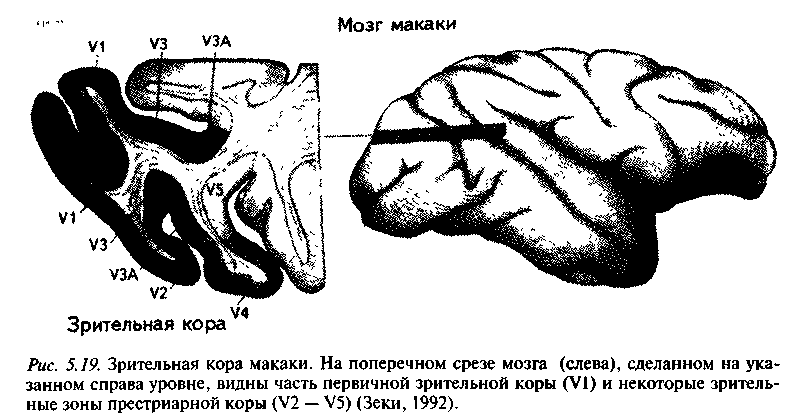

Это позволило выдвинуть концепцию функциональной организации зрительной коры, согласно которой цвет, форма, движение и, возможно, другие атрибуты видимого мира обрабатываются мозгом по отдельности. Поскольку сигналы к таким специализированным зонам идут преимущественно от VI, можно предположить, что у этой области, как и у V2, принимающей сигналы от VI и связанной с теми же зонами, также должна быть функциональная специализация. Возможно, две эти области действуют как своего рода почтовые отделения, сортирующие сигналы в зависимости от места назначения (Зеки, 1992).
Новые методы исследований подтвердили это предположение и позволили выявить специфические нервные пути, распространяющиеся от VI по всей престриарной коре. В одном из таких исследований оценивали кровоток у людей с нормальным зрением методом позитронно-эмиссионной томографии. Испытуемые рассматривали мозаику Э. Лэнда в стиле П. Мондри-ана (абстрактная картина из цветных прямоугольников без конкретных различимых объектов). Э. Лэнд — психофизик, разработавший этот тест, П. Монд-риан (1872-1944), голландский живописец, создатель неопластицизма — абстрактных композиций из прямоугольников, окрашенных в основные цвета спектра (рис. 5.20).
Было показано, что значительнее всего кровоток усиливался в веретенообразной извилине. Ее обозначили как зона V4 человека. Другие результаты были получены при рассматривании человеком черно-белого изображения движущихся прямоугольников: максимальная интенсивность кровотока зарегистрирована в более латеральной области — зоне V5 человека. При обоих типах стимуляции в первичной зрительной коре — зоне VI — также заметно усиливался кровоток. Вероятно, у человека, как и у обезьян, имен-
136
Рис. 5.20. Различные типы объектов стимулируют неодинаковые участки зрительной коры. Как показывает изучение кровотока в отдельных участках мозга, яркое полотно в стиле Мон-дриана сильно активирует зону V4, а движущиеся черно-белые фигуры — зону V5. Оба типа объектов приводят также к активации зон VI и V2 — функционально менее специализированных и распределяющих сигналы по другим корковым зонам (Зеки, 1992).
137

но первичная зрительная кора участвует в распределении сигналов по другим участкам коры.
Стриарная кора разделена приблизительно на 2500 колонок (модулей), каждая размером приблизительно 0,5 х 0,7 мм, и содержит примерно 150 000 нейронов. Ключ к работе распределительной системы зон заключается в их структурно-функциональной организации. Область VI необычно богата клеточными слоями. Если использовать метод окрашивания на присутствие цитохромоксидазы, ключевого фермента клеточного дыхания, то можно выявить клетки с высокой метаболической активностью (Wonh-Riley, 1978). По результатам окрашивания в стриарной коре выявляются метаболические колонки клеток. Если рассматривать их на срезе, параллельном поверхности коры, они выглядят интенсивно окрашенными пятнами или пузырьками, отделенными друг от друга более светлыми областями. М. Ливингстон и Д. Хью-
138
бель обнаружили, что клетки, избирательно реагирующие на определенную длину волны, сконцентрированы именно в таких пузырьках, а клетки, воспринимающие форму, — в областях между пузырьками (Livingstone, Hubel, 1987) (рис. 5.21).
Колонки особенно заметны во втором и третьем слоях зоны VI, получающих сигналы от парвоцеллюлярных слоев наружного коленчатого тела. Клетки этих его частей мощно и устойчиво реагируют на зрительные стимулы, причем многие из них — на цветовые.
Особый набор структур можно различить в слое 4В зоны VI, получающем сигналы от магноцеллюлярных слоев наружного коленчатого тела, клетки которого реагируют на движение кратковременно и в основном безразличны к цвету. От 4В отходят пути к зонам V5 и V3. Клетки 4В, связанные с V5, собраны в мелкие группы, окруженные клетками, сообщающимися с другими зрительными зонами. Организация слоя 4В зоны VI свидетельствует о специализации определенных ее частей на восприятии движения и о том, что они отделены от областей с другими функциями. Зона V2, как и VI, обладает особой метаболической архитектоникой, однако она образована широкими и узкими полосами, отделенными друг от друга менее интенсивно окрашиваемыми межполосными областями. Клетки, избирательно реагирующие на определенную длину волны, сосредоточены в узких полосах, нейроны, меняющие свою активность при изменении направления движения, — в широких, а клетки, чувствительные к форме, распределены как в широких полосах, так и между ними.
Таким образом, в зонах VI и V2 как бы находятся ячейки, где собираются различные сигналы перед передачей их в специализированные зрительные зоны. Клеткам этих ячеек соответствуют маленькие рецептивные поля, т. е. они реагируют только на стимулы, попадающие на ограниченные участки сетчатки. Кроме того, они регистрируют информацию только о специфических параметрах окружающего мира в пределах своего рецептивного поля. Они осуществляют как бы пофрагментный анализ всего зрительного поля (Зеки, 1992).
По-видимому, существует четыре параллельные системы, соответствующие различным аспектам зрения: одна связана с анализом движения, вторая — с анализом цвета, и две оставшиеся воспринимают формы объектов. Наиболее отличаются друг от друга системы, воспринимающие движение и цвет. За восприятие движения отвечает престриарная зона V5. Сигналы сюда идут от сетчатки через магноцеллюлярные слои наружного коленчатого тела и слои 4В зоны VI, откуда они попадают к месту назначения непосредственно, а также через широкие полосы V2. Цветовая система зависит от зоны V4; импульсы к ней поступают непосредственно или через узкие полосы V2 от пузырьков зоны VI, а к ним проходят от сетчатки через парвоцеллюлярные слои наружного коленчатого тела.
Избирательность реакций детекторов цвета на цветовые стимулы подготавливается на нескольких уровнях. На уровне рецепторов спектральное излучение вызывает комбинацию реакций трех типов колбочек. Таким образом, цветовой стимул кодируется вектором возбуждения рецепторов. На уровне горизонтальных клеток достигается переход к трем типам нейронов, оппонентные характеристики которых образуют ортогональный базис. На
139
уровне биполярных клеток сетчатки из возбуждений трех типов колбочек и трех типов горизонтальных клеток образуется четыре основных типа нейронов: красно-зеленый, сине-зеленый, яркостный и темновой. Следует специально подчеркнуть роль темнового нейрона, активного в темноте. С его участием происходит переход к вектору возбуждения постоянной длины, но уже в четырехмерном пространстве. Это осуществляется благодаря тому, что из активности темнового нейрона вычитаются возбуждения всех остальных нейронов так, что сумма возбуждений всех нейронов остается постоянной. Этот принцип кодирования цвета вектором возбуждения постоянной длины сохраняется и на уровне наружного коленчатого тела перед входом в зрительную кору. Четыре типа цветокодирующих нейронов наружного коленчатого тела питают цветоселективные детекторы коры и могут поэтому быть названы предетекторами. В зрительной коре происходит переход четырех типов нейронов-предетекторов к цветовым детекторам, реагирующим на отдельные оттенки. Это достигается тем, что цветовые детекторы обладают различными комбинациями весов своих синапсов (векторов синаптических связей), что позволяет детекторам избирательно реагировать только на определенную комбинацию возбуждений предетекторов (вектор возбуждения предетекторов). Совокупность цветоселективных детекторов образует детекторную карту, на которой отдельные цвета представлены точками.
Вектор возбуждения предетекторов характеризуется постоянной длиной, и его конец остается поэтому на сферической поверхности в четырехмерном пространстве, которую принято называть гиперсферой. Цветоселективные детекторы занимают отдельные участки цветовой гиперсферы. При изменении спектра излучения на входе вектор возбуждения предетекторов изменяет свою ориентацию, его конец перемещается на другой конец гиперсферы, возбуждая там другой детектор. Возбуждение цветоселективного детектора вызывает соответствующее ощущение цвета. Такая сферическая модель цветового зрения позволяет непротиворечиво интегрировать нейрофизиологические и психофизиологические данные в рамках единой нейронауки (Соколов, 2000).
Возбуждения четырех типов цветовых предетекторов образуют декартовы координаты цветового пространства. Сферические координаты того же цветового стимула (три угла гиперсферы) соответствуют субъективным шкалам: цветовому тону, светлоте, насыщенности. Сферическая модель цветового зрения позволяет по реакциям нейронов-предетекторов (декартовым координатам) найти субъективные шкалы (сферические координаты), а по их значениям представить возбуждения оппонентных нейронов-предетекторов (Хьюбел, 1990). Кроме того, сферическая модель, основанная на векторном кодировании, позволяет охарактеризовать субъективные различия между цветами как расстояния между концами представляющих их векторов возбуждения. Вычисление цветовых различий реализуется в фазических нейронах и проявляется в амплитуде негативного компонента вызванного потенциала N87, возникающего при замене одного цвета другим. Амплитуда N87 в высокой степени положительно коррелирует с субъективным различием между заменяемыми цветами.
Универсальность сферической модели цвета заключается в том, что она воспроизводит результаты выработки условных рефлексов на цветовые сти-
мулы у животных трихроматов. Цветовые названия, избирательно возникающие у человека при категоризации цветовых стимулов, также включены в сферическую модель. Аналогичная сферическая структура проявляется в зрительном анализаторе восприятия движения, а также в восприятии ориентации и пространственной частоты линий (Соколов, 2000).
Из двух формовоспринимающих систем одна связана с цветовой, а вторая от нее не зависит. Первая находится в зоне V4 и получает сигналы от пар-воцеллюлярных слоев наружного коленчатого тела через межпузырьковую часть VI и межполосную V2. Вторая локализована в зоне V3 и больше связана с динамическими формами, т. е. очертаниями движущихся объектов. К ней сигналы идут от магноцеллюлярных слоев наружного коленчатого тела через слой 4В зоны VI, а затем напрямую или через широкие полосы V2. Анатомия этих областей позволяет им сообщаться друг с другом.
Точечное повреждение этих зон ведет к различным видам патологии. Нарушения в зоне V4 лежат в основе ахроматопсии, при которой больные видят лишь оттенки серого цвета, не только не воспринимая, но и не осознавая других цветов. Нарушения в зоне V5 приводят к акинетопсии: больные не видят и не осознают движения в окружающем мире. Объекты внешнего мира исчезают из их восприятия и осознания, если начинают двигаться, и вновь становятся доступными при окончании движения. Подобные виды патологии свидетельствуют о том, что отдельные зоны зрительной коры самостоятельно перерабатывают информацию.
Интеграция зрительных ощущений происходит не в единой высшей области восприятия, а осуществляется посредством обширных прямых и обратных связей между отдельными специализированными областями на всех уровнях.

Рис. 5.22. Треугольник Каниша образован иллюзорными линиями. Нормальная зрительная кора воспринимает его, несмотря на их отсутствие на рисунке. Такого рода иллюзии показывают, что кора должна разрешать конфликты между различными функциональными зонами (Зеки, 1992).
Обратные связи необходимы для разрешения возможных конфликтов между клетками, реагирующими на одни и те же стимулы, но имеющими различные возможности в их обработке. Так, реакции клеток зон VI и V2 демонстрируют конфликт в иллюзии “треугольник Каниша” (рис. 5.22). Наблюдатель в этой иллюзии видит треугольник среди предъявляемых фигур, хотя линии, образующие его, не завершены, и мозг сам воссоздает недостающие отрезки. Р. фон дер Хейдт и Э. Петерханс (Зеки, 1992) показали, что клетки первичной зрительной коры, избирательно реагирующие на форму, не отвечают на эту иллюзию, тогда как нейроны V2, получающие импульсы от нее, уже чувствительны к ней. Для разрешения этого конфликта необходима обратная связь от V2 к VI.
140

Рис 523 Схематическое изображение роли “сенситивного” периода в формировании долговременных пластических перестроек синапсов на гностической единице Gr под влиянием активации А — активация, Gt , G, Gl+1 Gi+2— гностические единицы, S, St Sk — 'стимулы, достигающие гностических единиц (Соколов, 1996)
Анализ современных данных позволил сформулировать теорию многоступенчатой интеграции зрительной информации (Зеки, 1992). В соответствии с ней интеграция протекает не в один этап благодаря конвергенции сигналов в некоторой высшей точке и не откладывается до тех пор, пока все зрительные зоны завершат анализ информации. Она представляет собой процесс одновременного восприятия и осознания окружающего мира и требует существования обратных связей между всеми специализированными зонами. Эта теория подтверждается многочисленными экспериментами.
Формирование восприятия целостного зрительного образа, согласно представлениям Е.Н. Соколова (1996), связано с конкретным вектором. Вектор представляет собой комбинацию возбуждений в ансамбле нейронов. Компонентами такого вектора являются возбуждения нейронов-детекторов отдельных признаков зрительного образа. Они конвергируют на нейронах более высокого порядка, которые обладают способностью реагировать на сложные изображения (например, изображение лица). Такие нейроны называются гностическими единицами. Объединение нейронов-детекторов отдельных признаков происходит посредством включения их в иерархически организованную нейронную сеть. Каждому зрительному образу соответствуют свои гностические единицы.
Формирование гностических единиц происходит в такой последовательности (Соколов, 1996). В передней вентральной височной коре имеется пул резервных нейронов, слабо реагирующих на стимулы. Под влиянием сигнала новизны из гиппокампа они активируются, причем включается механизм пластических перестроек в синапсах. В это время сигнал, поступающий от детекторов к резервному нейрону, улучшает синаптическую связь между ними. Вследствие этого нейрон селективно настраивается на восприятие такого стимула. По окончании сенситивного периода процесс обучения прекращается и нейрон утрачивает способность формировать новые связи (рис. 5.23).