

Репродуктивный цикл
Репродуктивный цикл у женщин называется менструальным, в то время как у других млекопитающих есть эстралъный цикл, или эструс, не имеющий месячной периодичности. В менструальном цикле регулярно происходит рост и уменьшение стенки матки. Цикл состоит из последовательности событий, которые контролируются гормонами гипофиза и яичника.
Гонадотропин гипоталамуса через гонадотропный гормон гипофиза в начале цикла способствует росту фолликула в яичнике. Это проявляется в том, что вокруг яйцеклетки разрастаются эпителиальные клетки. Обычно в одном цикле развивается одна яйцеклетка, хотя иногда возможно развитие одновременно двух и более яйцеклеток, что может привести к рождению дизиготных близнецов. Созревая, фолликул секретирует гормон эстрадиол, вызывающий утолщение стенки матки, что подготавливает ее к имплантации яйцеклетки (рис. 17.5).
Возрастающая секреция эстрадиола является сигналом выброса рели-зинг-фактора лютеинизирующего гормона из гипоталамуса. Это вещество вызывает выделение лютеинизирующего гормона из гипофиза, обусловливающего овуляцию, т. е. выход яйцеклетки из фолликула. Фолликул превращается в лютеинизирующее тело (желтое тело), которое вырабатывает прогестерон и эстрадиол. Прогестерон обеспечивает сохранение беременности. Он поддерживает утолщение матки и подавляет рост других фолликулов.
Созревшая яйцеклетка после овуляции движется по фаллопиевым тру-
387
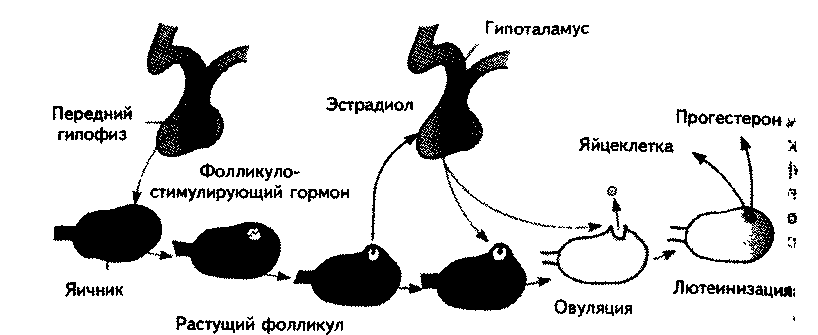
Рис. 17.5. Нейроэндокринный контроль менструального цикла (Carlson, 1992).
бам в сторону матки. Если она за это время оплодотворяется, то, непрерывно делясь, через несколько дней внедряется в стенку матки. Если встречи со сперматозоидом не происходит, то имплантация в стенку матки не осуществляется. В этом случае желтое тело перестает продуцировать эстрадиол и прогестерон, что приводит к отслаиванию стенки матки, т. е. начинается менструация.
Нервный контроль сексуального поведения
Медиальная преоптическая область, расположенная в ростральном отделе гипоталамуса, непосредственно связана с мужским сексуальным поведением. Ее электрическое раздражение у крыс ведет к попыткам копуляции (половому акту) (Malsbury, 1971), что сопровождается нарастанием метаболической активности в ней (Oaknin e. а., 1989). Разрушение медиальной пре-оптической области приводит к исчезновению такого поведения (Heimer, Larsson, 1966/67). Исследования на обезьянах показали, что электрическое раздражение этого участка мозга может вызвать эрекцию пениса, эякуляцию или мастурбацию (MacLean, 1973).
Именно через медиальную преоптическую область гипоталамуса андро-гены осуществляют свою функцию, поскольку здесь в пять раз больше анд-рогеновых рецепторов у самцов, чем у самок (Roselli e. а., 1989), а ее размер в три-семь раз больше у самцов по сравнению с самками (Gorski e. а., 1978). В то же время величина этого участка уменьшается, если животное в пре-натальный период подвергалось стрессу (Anderson e. а., 1986).
Половые гормоны различным образом влияют на мозг животных разного пола. Обнаружено, что у самцов размер гиппокампа больше справа, тогда как у самок — слева (Diamond e. а., 1981).
Из медиальной преоптической области гипоталамуса аксоны направляются в тегментальную область среднего мозга (дорзальнее и медиальнее от
388
черной субстанции). Таким образом, эта область проявляет свое действие через моторные структуры среднего мозга.
У человека различия в размерах преоптической области между мужчинами и женщинами обнаруживаются только с четырехлетнего возраста (Swaab, Hofrnan, 1988). К сексуальному поведению у людей имеет отношение так-лсе височная доля мозга, поскольку ее дисфункция ведет к снижению сексуального желания. Например, эпилептические припадки, обусловленные повреждением височной области, сопровождаются снижением сексуального интереса (Blumer, 1975).
Сексуальное поведение женщин в большей мере связано с вентромеди-альным ядром гипоталамуса. Повреждение этого ядра ведет к отсутствию лордоза у самок крыс. Раздражение его электрическим током, так же, как и инъекция непосредственно в эту область эстрадиола и прогестерона, напротив, способствует стимуляции сексуального поведения (Pfaff, Sakuma, 1979). Свое влияние на поведение вентромедиальное ядро осуществляет через аксоны, направляющиеся в околоводопроводное серое вещество среднего мозга, а затем — в ретикулярную формацию и в спинной мозг (Carlson, 1992).
Электрическая стимуляция лобных долей у человека также вызывает оргазмоподобное переживание. Нервные центры, регулирующие оральные ответы, тесно связаны с центрами, регулирующими генитальные реакции: их низкочастотное раздражение вызывает сначала слюноотделение и жевательные движения, а примерно через минуту — эрекцию полового члена. Это объясняет тот факт, что у ребенка эрекция полового члена часто наблюдается при кормлении. П.Д. Маклин (MacLean, 1973) полагает, что связь оральных и генитальных реакций может опосредоваться обонянием, поскольку запахи являются обязательным компонентом как пищевого, так и сексуального поведения животных.