

2.2. Проблема индивидуальных различий ключевая проблема биологии продолжительности жизни
Продолжительность жизни особей одного вида, живущих в сходных условиях, колеблется в очень широких пределах. С одной стороны, жизнь может оборваться еще до рождения. По современным данным, такова участь 70—80Х оплодотворенных яйцеклеток человека, причем в большинстве случаев гибель происходит на столь ранней стадии, что обычно никем не замечается [Diamond, 1987]. С другой стороны, длительность жизни отдельных индивидуумов может значительно превышать соответствующие средние значения для всей популяции, причем абсолютной верхней границы длительности жизни, по-видимому, вообще не существует (к этому вопросу мы вернемся несколько позднее в разделе 4.2. данной книги) Но даже если отбросить все случаи гибели организма до рождения и факты удивительного долголетия, то все равно окажется, что индивидуальные различия по срокам жизни довольно велики. Например,в 1974—1978 гг. средняя продолжительность жизни мужчин в Швеции составляла 72.2 года[BefolkningsfOrandringar,1970— 1981]. При этом вероятность умереть в возрастном интервале 71— 73 года составляла менее 6Х. В других странах эта вероятность оказывается еще меньше. Можно, конечно,объяснить высокую вариабельность длительности жизни людей социальными и генетическими различиями между ними, не говоря уже о климатических и биогеохимических особенностях отдельных районов страны. Но та же картина наблюдается и в популяциях линейных лабораторных
20
животных. Так. например,в популяции 1415 самок лабораторныхдрозофиллинии 107[Pearl,Parker, 1924a] со средней продолжительностью жизни 48 дней в возрасте 43—49 дней погибло всего ИХ всех особей.
Одной из характеристик вариабельности длительности жизни может служить коэффициент ее вариации (отношение среднего квардратичногоотклонения к среднему арифметическому, выраженное в процентах). Расчет этих коэффициентов на примере лабораторных дрозофил показывает,что коэффициент вариации длительности жизни составляет обычно около 35Х, достигая иногда 65Х [Семенова, 1983].
Таким образом, факт значительной биологической вариабельности длительности жизни не вызывает сомнений и требует своего объяснения, а также количественного описания. Есть еще одно обстоятельство. выдвигающее эту проблему на первый план.
В отличие от большинства других количественных признаков (например. длины или температуры тела) продолжительность жизни организма можно измерить только один раз. Этот тривиальный факт имеет далеко идущие последствия. Длительность жизни, оказывается, в принципе невозможно изучать на организменномуровне, поскольку нельзя исследовать индивидуальную динамику этого признака и определить ошибку единичного наблюдения. Поэтому единственным способом изучения продолжительности жизни остается еепопуляционно-статистическоеисследование при варьировании генетических исредовыхфакторов. Но ведь такой подход в принципе не может дать ничего, кроме информации о распределении длительности жизни в изучаемой популяции и влиянии различных факторов на это распределение! Таким образом, проблема расшифровки данных о распределении длительности жизни является ключевой для всех дальнейших исследований.
Знание закона распределения продолжительности жизни оказывается полезным уже на самых первых этапах исследования. Действительно, результаты экспериментов по выживаемости, представленные рядом значений продолжительности жизни для каждого организма, слишком громоздки для их эффективного анализа, поэтому возникает необходимость в более компактной записи результатов эксперимента. Отчасти эта проблема решается путем построения таблиц смертности [Дубина, Разумович.1975; Коли, 19791.
Обычно таблица смертности составляется для группы одновременно родившихся индивидуумов (когорты) и описывает наблюдаемую картину смертности до того момента, когда умрет последний член этой группы. Такие таблицы смертности называются когортными и широко используются при изучении продолжительности жизни лабораторных и диких животных [Leslie,Ranson,1940;Miller,Thomas, 1958;Pearl,Parker, 1928a].
Рассмотрим метод построения когортнойтаблицы смертности более подробно[Chiang,1978]. Пусть численность когорты измеряется через одинаковые промежутки времени, равные я- Соответственно на
21
Таблица 1
Таблица смертности самцов имаго Drosophita melanogaster (рассчитано на основании данных [Pearl, Parker, 1924a])
|
Возрастной интервал (дни)
|
Число доживающих до возраста x
|
Число умерших в интервале (х, х + n)
|
Вероятность смерти в интервале (х,х+n)
|
Суммарное время жизни в интервале (х, х + n)
|
Суммарное время жизни в возрасте х и старше
|
Ожидаемая продолжительность жизни в возрасте х
|
|
(х, х+n)
|
lx
|
dx
|
qx
|
lx
|
Тх
|
eх
|
|
0—6 7—12
|
1407 1389
|
18 12
|
0,01279 0,00864
|
8388 8298
|
63657 55269
|
45,2 39,8
|
|
13—18 19—24
|
1377 1351
|
26 85
|
0,01888 0,06292
|
8184 7851
|
46971 38787
|
34,1 28,7
|
|
25—30 31—36
|
1266 1166
|
100 130
|
0,07899 0,11149
|
7296 6606
|
30936 23640
|
24,4 20,3
|
|
37—42 43-48
|
1036 848
|
188 173
|
0,18147 0,20401
|
5652 4569
|
17034 11382
|
16,4 13,4
|
|
49—54 55—60
|
675 469
|
206 285
|
0,30519 0,60768
|
3432 1959
|
6813 3381
|
10.1 7,2
|
|
61-66 67—72
|
184 101
|
83 62
|
0,45109 0,61386
|
855 420
|
1422 567
|
7,7 5,6
|
|
73—78 79—84
|
39 4
|
35 3
|
0,89744 0,75000
|
129 15
|
147 18
|
3,8 4,5
|
|
85+
|
1
|
1
|
1,00000
|
3
|
3
|
3,0
|
ту же величину будет каждый раз увеличиваться и возраст членов когорты. Основными величинами,входящими вкогортнуютаблицу смертности,являются/д— число доживших до возрастах; d^ —число умерших в интервале(х, х+я).
Между величинами ^и d^существует следующее соотношение:

Вероятность смерти q^в каждом возрастном интервале (х,х+и) можно получить делением d^на/.,:
![]()
Число живущих, или, точнее, суммарное время жизни, L^в возрастном интервале (х,х + п)обычно рассчитывается по формуле:
![]()
Для расчета ожидаемой продолжительности жизни необходимо сначала вычислить величины Г»для каждого возраста, которые представляют собой суммарное время жизни всех членов когорты в возрастехи старше:
![]()
где (о — начало последнего возрастного интервала. И наконец, ожидаемая продолжительность жизни еxрассчитывается следующим образом:
![]()
где x=0, 1, ..., ω.
В качестве примера когортнойтаблицы смертности можно привести таблицу смертности имаго Drosophila melanogaster(табл. 1). На рис. 1а.б, в, гприведены типичные зависимости показателей таблицы смертности от возраста.
Для построения таблиц смертности человека приведенным выше способом потребовался бы срок наблюдения, близкий к 100 годам, поэтому таблицы смертности людей обычно рассчитываются другим методом, который будет описан в разделе 3.1. данной книги.
К настоящему времени построено большое количество таблиц смертности не только лабораторных,но также домашних [Hickey, 1960] и диких [Caughley,1966; Deevey,1947] животных и даже растений[Solbrig,1980]. На составление этих таблиц затрачен и продолжает затрачиваться колоссальный труд. а их значение для исследования биологии продолжительности жизни трудно переоценить. По существу, эти таблицы являются единственным экспериментальным
23
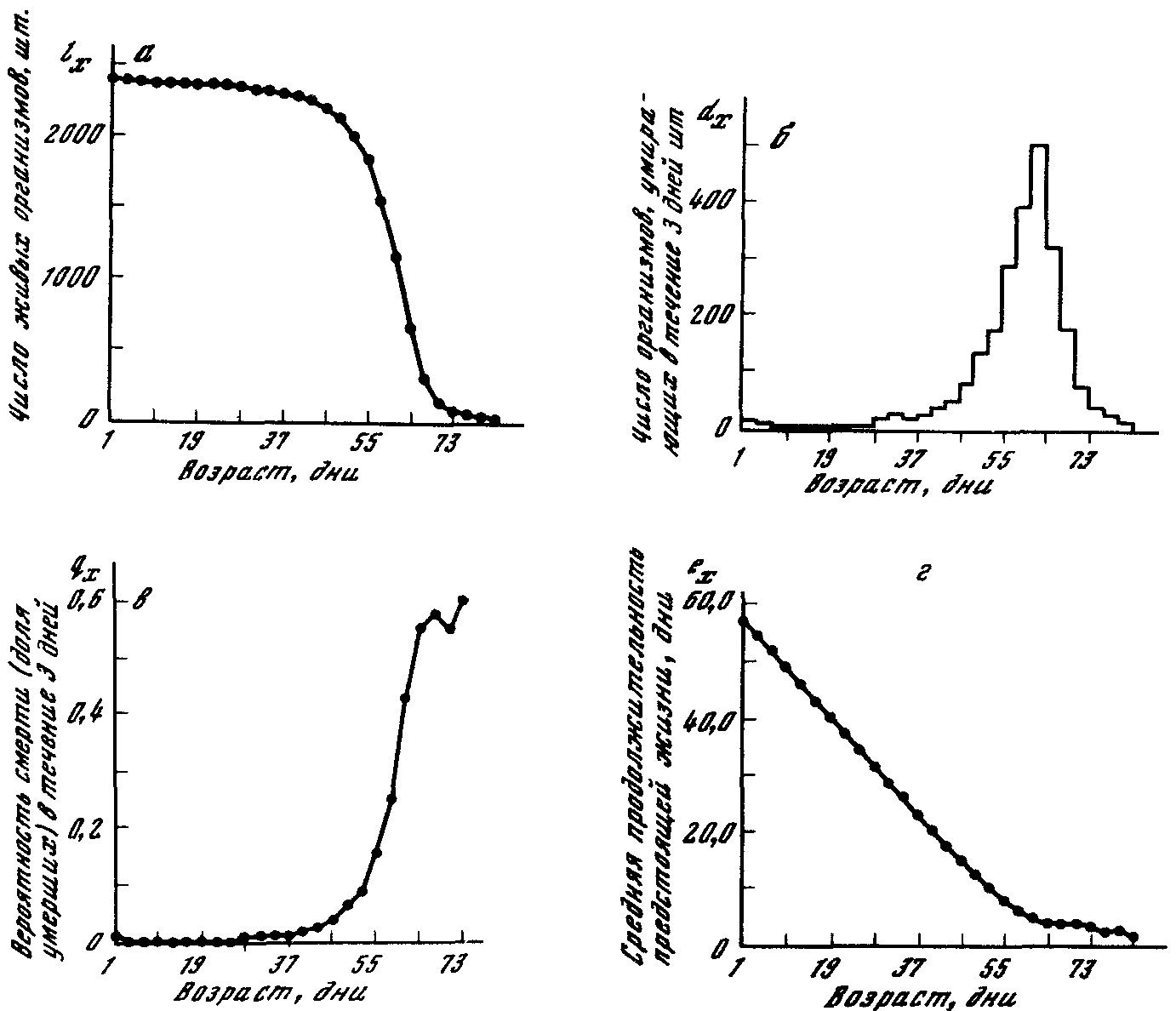
Рис I. Зависимость показателей таблицы смертности от возраста:
а. — кривая дожития (/д), б— кривая числа умерших (ау; в — вероятность смерти (ЧхУ- г~ средняя продолжительность предстоящей жизни (е^)
Рассчитано и построено на основании данных по выживаемости 2400 самок Drosophila melanogaster, опубликованных в работе [Hall. 1969]
источником для таких исследований. Между тем опубликованные таблицы смертности животных разбросаны по многим изданиям. посвященным самым разным проблемам: геронтологии, онкологии, экологии, радиобиологии и др. Далеко не во всех работах,содержащих анализ продолжительности жизни, приводятся собственно таблицы смертности. До сих пор нет исчерпывающей библиографии работ, в которых опубликованы таблицы смертности животных.
В связи с этим сведение воедино опубликованных таблиц смертности животных может оказать помощь в дальнейшей разработке проблемы биологии продолжительности жизни. В табл. 2 приводится перечень опубликованных таблиц смертности животных с указанием условий проведения эксперимента.
Однако подход, связанный с использованием таблиц смертности, все-таки далек от совершенства. В самом деле, с одной стороны. данные таблицы (особенно полные таблицы смертности человека) все-
24
Таблица 2
Перечень опубликованных таблиц продолжительности жизни животных (в таблице приведены лишь наиболее надежные данные, полученные для достаточно больших выборок)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
1. ТИП ASCHELMINTHES (ПЕРВИЧНОПОЛОСТНЫЕ ЧЕРВИ)
| ||
|
< 1.1. Класс Nematoda (круглые черви)
| ||
|
Caenorhabditis elegans
|
1
|
Findei» et al., 1983
|
|
12. Класс Rotatoria(KonoepaTKB)
| ||
|
llydantyna senta миктическиеи амиктические самки
|
3
|
Ferris, 1932
|
|
Proatessordida Proales sordida Proales declpiens
|
5 8 1
|
Jennings, Lynch, 1928a Jennings, Lynch, 1928b Noyes, 1922
|
|
2. ТИП MOLLUSCA (МОЛЛЮСКИ)
| ||
|
2.1. Класс Gastropoda (брюхоногие моллюски)
| ||
|
2.1.1. Подкласс Pulmonata (легочные моллюски)
| ||
|
Biomphalaria pfeijferi
|
|
|
|
различная температура Bulinus globosus
|
6
|
DeKock, van Eeden, 1981
|
|
различная температура Bulinus tropiciis влияние размера попул. различная температура . содержания .
|
4 2 20 3
|
Shiff, 1964 Bariow, Muench, 1951 De Kock, van Eeden, 1976 De Kock, 1985
|
|
различная температура . содержания .
|
6
|
De Kock, van Eeden, 1985
|
|
последовательные генерации Planorbis boissyi Lymnea columella (прудовики) Lymnea natalensis
|
10 2 2
|
De Kock et al., 1986 Bariow.Muench.l951 Winsor, Winsor, 1935
|
|
различная температура . содержания .
|
3
|
De Kock, 1985
|
|
различная температура . содержания
|
6
|
DeKock.vanEeden.l985
|
|
Agriolimax agreslis (полевой слизень)
|
4
|
Szabo,Szabo,1929
|
|
3. ТИП ARTHROPODA (ЧЛЕНИСТОНОГИЕ)
| ||
|
3.1. Класс Crustacea (ракообразные)
| ||
|
Daphnia pulex (обыкновенная дафния) различная плотность популяции Daphnia magna
|
7
|
Francketal.,1957
|
|
различная температура
|
8
|
MacAnhur, Bafflie, 1929
|
25
Таблица 2 (продолжение)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
3.1. Класс Crustacea (ракообразные)
| ||
|
Balanus balanoides (обычный морской желудь)
|
1
|
Bodenheimer, 1958
|
|
Arthemia salina (артемия) облучение
|
2
|
Suyama, Iwasaki, 1976
|
|
3.2. Класс Arachnida (паукообразные)
| ||
|
Latrodectus maclans (паук черная вдова)
|
2
|
Deevey, Deevey, 1945
|
|
Paruroctonus mesaensis (скорпион)
|
2
|
PoUs, Parley, 1980
|
|
3.3. Класс Insecta (насекомые)
| ||
|
3.3.1. Отряд Lepidoptera (чешуекрылые: бабочки)
| ||
|
Fwnea crassiorelta Samia cercopia Cullosarma promelhea Tropaealuna Bmnbyx mori (шелкопряд) Acrobasis caryae (чехлоноска) Carpocapsa pomomlla
|
1 12 2 2 4 8 8
|
Matlhes, 1951 Rau, Rau, 1912 Rau, Rau,1912 Rau, Rau, 1912 Алпатов, Гордеенко, 1932 Pearl, Miner, 1936 McAnhur, Baillie, 1932
|
|
3.3.2. Отряд Coleoptera (жесткокрылые: жуки)
| ||
|
Rhizopertha dominica (зерновой ТОЧИЛЬЩИК) Cahndra oryzae (рисовый долгоносик)
|
2
|
Birch,1953
|
|
различная влажность, температура и корм
|
10
|
Birch, 1953
|
|
Trogodenna versicolor (жук-кожеед) Tribolium caslaneum (каштановый хрущак)
|
4 8
|
Davis, 1945 Meitzetal.,1965
|
|
Tribolium madens (черный мучной хрущак)
|
2
|
Park, 1945
|
|
Tribolium confusum (малый мучной хрущак), облучение
|
3
|
Cork, 1957
|
|
—-—
|
8 2
|
Mertz et al., 1965 Pearl, Miner, 1941
|
|
3.3.3. Отряд Diptera (двукрылые: мухи, комары и москиты)
| ||
|
Phormia terrae novae (мухиформии)
|
1
|
Collatz, Wilps, 1983
|
|
Calliphora erythrocephala (синяя мясная муха)
|
2
|
Feldman-Muhsam, Muhsam, 1945
|
|
Musca vicina (южная комнатная муха)
|
2
|
Feldman-Muhsani, Muhsam, 1945
|
|
Musca domeslica (комнатная муха)
|
2
|
Rockstein, Liebennan, 1959
|
|
различный корм
|
10
|
Rockstein, 1957
|
26
Таблица 2 (продолжение)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
3.3.3. Отряд Diptera (двукрылые: мухи, комары и москиты)
| ||
|
Similium venustum Drosophila psendoobscura Drosophila melanogaster
|
6 15
|
Davies, 1953 Vetukhiv, 1957
|
|
различная плотность популяции в личиночной стадии
|
12
|
Miller, Thomas, 1958
|
|
различная температура линия Canton-S дикая линия, а также мутанты cinnabar, curled, vestigial, while, yellew (асептические условия)
|
9 4 30
|
Loeb.Nonhrop.l917 Hall, 1969 Steinfeld, 1929
|
|
линии Old Falmouth, Quintuple и их pi, F; и рз — гибриды
|
35
|
Peari et al., 1923
|
|
линия Old Falmouth, различная плотность популяции
|
26
|
Peart el al., 1927
|
|
линии Old Falmouth, New Falmouth, Sepia, Eagle Point и Quintuple
|
8
|
Pearl, Parker, 1927
|
|
линии Old Falmouth и New Falmouth
|
14
|
Pearl, Parker,1922a
|
|
обработка эфиром различная плотность популяции
|
11 50
|
Pearl, Parker, 1922b Pearl, Paiker, 1922с
|
|
содержание при различной вентиляции воздуха
|
9
|
Pearl, Paiker, 1922d
|
|
линии Old Falmouth и vestigial
|
4
|
Pearl, Parker, 1924a
|
|
полное отсутствие пищи, различная плотность популяции
|
12
|
Pearl, Parker, 1924b
|
|
дикая линия и мутанты vestigial, nicked, notched-b-pr и их гибриды
|
14
|
Luers, 1935
|
|
дикая линия и мутант ebony, полное отсутствие пищи
|
4
|
Greiff, 1940
|
|
Самцы, самки, триплондные самки и ннтерсексы
|
6
|
Gowen, 1931
|
|
Мутанты vestigial, black, speck, arc, purple и комбинации перечисленных мутаций
|
55
|
Gonzales, 1923
|
|
различная температура во время личиночной и имагинальной стадий
|
12
|
Alpatov, Pearl, 1929
|
27
Таблица 2 (продолжение)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
различное питание во время личиночной и имагинальной стадий
|
12
|
Alpatov, 1930
|
|
Anopheles dirus (комар) Aedes aegyptl (желтолихорадочный комар)
|
6 2
|
Klein et al., 1987 Kershawetal., 1953
|
|
3.3.4. Отряд Hymenoptera (перепончатокрылые: муравьи, осы, пчелы)
| ||
|
Habrobraconjuglandis (оса-наездник)
|
4
|
Georgiana, 1949
|
|
Bracon kirkpatriclu
|
1
|
Sivaprakasam et al., 1985
|
|
3.3.5. Отряд Blalloptera (таракановые)
| ||
|
Blatla orientalis (черный таракан)
|
2
|
Rau,1924
|
|
3.3.6. Отряд Homoptera (равнокрылые)
| ||
|
Therioaphis maculala (пятнистая люцерновая тля) различная температура
|
2
|
Messenger, 1964
|
|
Fiorinia Iheae
|
2
|
Munir, Sailer, 1985
|
|
3.3.7. Отряд Anoplura (вши)
| ||
|
Pediculus humanus (головная вошь)
|
2
|
Evans, Smith, 1952
|
|
4. ТИП CIIORDATA (ХОРДОВЫЕ)
| ||
|
4.1. Подтип Urochordata (оболочники)
| ||
|
4.1.1. Класс Ascidiae (асцидии)
| ||
|
Ascidia nigra
|
1
|
Goodbody,1962
|
|
4.2. Подтип Vertebrata (позвоночные)
| ||
|
4.2.1. НадклассРisces (рыбы)
| ||
|
Lebisles reliculatus (гуппи) Oryzias talipes (японская медака) облучение Poecilopsis monacha Cynolebias bellottli (аргентинская цинолебия)
|
14 1 8 2 2
|
Comfort, 1963 Egami, Etoh, 1969 Egami, Etoh, 1973 Moore, 1985 Uu,Walfoid,1969
|
|
4.2.2. Надкласс Tetrapoda (четвероногие)
| ||
|
4.2.2.1. Класс Amphibia (амфибии: земноводные)
| ||
|
Desmognalhus fuscus (темная саламандра)
|
12
|
Danstedt, 1975
|
28
Таблица 2 (продолжение)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
4.2.2.2. Класс Reptilia (рептилии: пресмыкающиеся)
| ||
|
Via slansbwiana (пятннстобокая игуана)
|
1
|
Tinkle, 1967
|
|
Sceloporus virgalus Sceloporus undulatus (заборная игуана)
|
2 1
|
Vinegar, 1975 Ferguson et al., 1980
|
|
Chrysemys plcta (расписная черепаха)
|
2
|
Wabur. 1975
|
|
Opheodrys aeslivus
|
1
|
Plummer, 1985
|
|
4.2.2.3. Класс Aves (птицы)
| ||
|
Callus gallus (домашняя курица)
|
1
|
Gardner, Hurst, 1933
|
|
Lonchura striata (бенгальский вьюрок)
|
2
|
Eisner, 1967
|
|
Megadyptes adeliae (пингвин Адели)
|
1
|
Ainley, DeMaster, 1980
|
|
Megddyptes antipodes (ПИНГВИН)
|
1
|
Richdale, 1957
|
|
Fringilla coelebs (зяблик)
|
2
|
Паевский,1985
|
|
Slumus vulgaris (обыкновенный скворец) ErUhacus rubeculs (обыкновенная зарянка)
|
1 1
|
Coulson. 1960 Паевский.1985 Паевский,1985
|
|
Corms nwnedula (обыкновенная галка)
|
2
|
Beklova, Pikula, 1985
|
|
Anseranser (серый гусь)
|
1
|
—
|
|
Cygnus olor (лебедь-шипун)
|
1
|
-"-
|
|
Anas querquedula (чирок-трескунок)
|
1
|
-"-
|
|
Anas strepera (серая утка)
|
1
|
—"—
|
|
Anas clypeala (широконоска)
|
1
|
-"-
|
|
Aylhyaferinar (красноголовый нырок)
|
1
|
—"—
|
|
Accipiter genlilis (тетеревятник)
|
1
|
—"—
|
|
BiMeo buteo (канюк)
|
1
|
-"-
|
|
Gallinula chloropus (камышница)
|
1
|
-"-
|
|
Gallinago gallinago (обыкновенный бекас)
|
1
|
-"-
|
29
Таблица 2 (продолжение)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
4.2.2.3. Класс Aves (птицы)
| ||
|
Columbia palwnbus (вяхирь) Athene noctua (домовый сыч) Aria otus (ушастая сова) Corvus corone cornix
|
2 1 1 1
|
-"-
|
|
(серая ворона) Pica pica (сорока)
|
1 1 1
|
Beklova, Pikula, 1985 Holyoak, 1971 Tatter, 1986
|
|
Garrulus glandarius (сойка) Actilis hypoleucos (перевозчик)
|
1 1
|
Beklova, TOcula, 1985 Beklova, Kkula, 1985 -'-
|
|
Nycticorax nyclicorax
|
1
|
|
|
(кваква), Anatinae (утки) Striginae (совы) Accipitridae
|
1 1 1
|
-•-
|
|
(семейство ястребиных) Anas platyrhync has (кряква) Accipiter nisus (перепелятник)
|
3 1
|
-"-
|
|
Fulicaatra (лысуха) Strix aluco (обыкновенная неясыть)
|
1 1
|
-"-
|
|
Corvusfrugilegus (грач)
|
2
|
—"
|
|
4.2.2.4. Класс Mammalia (млекопитающие)
| ||
|
Отряд Insectivora (насекомоядные)
| ||
|
Blarina brevicauda (короткохвостая землеройка)
|
1
|
Blus, 1971
|
|
Отряд Chiroptera (рукокрылые: летучие мыши)
| ||
|
Myotis aaubentoni (водяная ночница)
|
1
|
Bezenetal.,1960
|
|
Myotis mustacinus (усатая ночница) Myotis emarginalus (трехцветная ночница)
|
1 1 1
|
Sluiteretal.,1956 Bezen et al., 1960
|
|
Rhinolophus hipposideros (малый подковонос)
|
1
|
-"-
|
|
Отряд Rodenlia (грызуны)
| ||
|
Microlus arvalis (полевка обыкновенная)
|
1
|
Фенюк. Шейкина, 1940
|
|
MicrMus agreslic (темная полевка)
|
1
|
Leslie, Ranson, 1940
|
|
Microtus orcadensis (оркнейская полевка)
|
2
|
Leslie et al., 1955
|
30
Таблица 2 (продолжение)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
Отряд Rodenlia (грызуны)
| ||
|
Ochotona princeps (американская пищуха)
|
1
|
Millar, Zwickel, 1972
|
|
Tapus princeps (западный полутушканчик)
|
1
|
Falk, MiUar, 1987
|
|
Spennophilus beldingi (суслик Белдинга) Spennophilus columbianus (колумбийский суслик)
|
2 1 6
|
Shennan, Morton, 1984 Barkalov et al., 1970 Zammuto, 1987
|
|
Cricetus auratus (сирийский хомячок) Rallus rattus (черная крыса)
|
2 4
|
Poiley, 1957 Fortner, 1961
|
|
линия Albany линия Wistar линия Wistar линия Wistar, упражнения Сингенные парабионты различного возраста
|
1 2 20 4 2
|
Wright et al., 1940 Weisner,Sheard,1935 Schlettwein-Gsell, 1970 Goodrick, 1980 Ludwig, Elashoff, 1972
|
|
линия Wistar линия Sprague-Dawley Линии Copenhagen, Fisher и их гибриды
|
2 1 22
|
Gilbert, Cillman, 1958 Durbinetal., 1966 Dunning, Curtis, 1946
|
|
линия Wistar линия Copenhagen, Fisher, August
|
1 10
|
Boonnan, Hollander, 1973 Curtis etal., 1931
|
|
Различные диеты Влияние экстрогенов Линия Wistar, действие прокаина
|
2 5 6
|
Ковтун.1976 Ross, 1961 Eisen, 1942 Asian et al., 1965
|
|
Raltus norvegicus (серая крыса, пасюк)
|
1
|
Leslie et al., 1952
|
|
Raltus (Mastomys) nalalensis Mus musculus (домовая мышь)
|
1 2
|
Oliff, 1953 Greenwood, 1928
|
|
облучение линия СЗН
|
4 1 2
|
Storer, 1965 Bittner, 1935b Andervont, McEleney, 1941
|
|
линия С линия C3Hf облучение линии DBA, C57BL и их fi и F^ — гибриды
|
2 3 3 12
|
Andervont, 1945 Vellisto, Bang, 1968 Storer, 1962 Murray, Little, 1935
|
|
линия DBA линия DBA н А линия DBA и C57BL и их гибриды
|
1 6 16
|
Murray,1934 Murray, Hoffman, 1941 Murray, HoUinan, 1939
|
|
линия C57BL
|
3
|
Little et al., 1939
|
31
Таблица 2 (продолжение)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
nfus musculus
|
|
|
|
линия SAS/4, облучение ор&льные контрацептивы гибриды А х DBA линия albino
|
8 5 1 1 3
|
Lindop, Rotblat, 1961 Garg et al„ 1970 Bitlner, 1936 Bittner, 1935a Bittner, 1937
|
|
линия А, различные диеты линия CC57BR
|
2 2 4 2
|
Bittier, 1935с Медведев,1958 Гельштейн, Дядькова, 1955 Медведев, Ольховская, 1974
|
|
линия CC57W линия CC57W. CC57BR линия СзНА линия DBR
|
2 8 3 2
|
Медведев, Ольховская, 1974 Медведев,1961 Малюгина, Прокофьева, 1957 Дядькова, Лотош, 1962
|
|
облучение
|
5 10
|
Thomson et al., 1985a Thomson et al., 1985b
|
|
быстрые нейтроны и уоблучение
|
24
|
Thomson, Grahn, 1988
|
|
линия Сз-black линия RF
|
1 1
|
Дядькова. Медведев, 1956 Yuhas, Clapp, 1972
|
|
Отряд Carnivora (хищные)
| ||
|
Canisfamiliaris (домашняя собака)
|
|
|
|
ирландские волкодавы пекинская болонка кокер-спаниэль английский мастифф биглн,облучение Cams lupus (волк обыкновенный)
|
2 1 1 1 3 1
|
Comfort, 1956 Comfort, 1960 Andersen, Rosenblatt, 1969 Comfort, 1957
|
|
Отряд Pinnipedia (ластоногие)
| ||
|
Halichoerus grypus (серый тюлень)
|
1
|
Hewer, 1964
|
|
Отряд Artiodaclyla (парнокопытные)
| ||
|
Capreolus capreolus (косуля) Odocoileus hemionus (чернохвостый олень)
|
2 2
|
Andersen, 1953 Taber,Dasmami,l958
|
|
Rangifor arcticus arclicus (арктический карибу)
|
1
|
Banfield, 1955
|
|
Hemilragus jemlalucus (тар) Alces alces (лось) Ammolragus lerv'ia (гривистый баран)
|
1 1 1
|
Caughley, 1966 Язан.19Б1 Comfort, 1957
|
|
Ovi? musimon (муфлон) Ov'is dalli (горный баран) Ovis aries (домашняя овца)
|
1 2 1
|
Comfort. 1957 Deevey,1947 Hickey, 1960
|
32
ТаблицаZ (окончание)
|
Биологический вид
|
Число таблиц
|
Источник данных
|
|
Отряд Artiodactyla (парнокопытные)
| ||
|
Syncerus coffer (африканский буйвол)
|
1
|
Sinslair, 1974
|
|
Boselaphus tragocamelus (антилопанильгау)
|
1
|
Comfort, 1957
|
|
OTp»nPerissodactyla(»empnoKOllvn»lde)
| ||
|
Diceros bicornis (черный носорог) Equus caballus (домашняя лошадь) Арабские скакуны
|
1 2 1
|
Goddari.1970 Comfort, 1958 Comfort, 1961
|
такиостаются слишком громоздкими. Например, полная демографическая таблица дожития, в которой приводятся значения показателей за каждый год возраста,содержит не менее 80 пар значений переменных с общим объемом массива данных свыше 500 значащих цифр[Keyfitz,1982].
С другой стороны, при построении подобных таблиц происходит некоторая потеря информации, связанная с группировкой данных по дискретным возрастным интервалам. Эта проблема могла быть решена, если бы был известен закон распределения продолжительности жизни. Например, если бы распределение организмов по срокам их жизни следовало нормальному закону,то любой сколь угодно большой массив данных о выживаемости можно было бы записать всего лишь парой чисел: величинами средней продолжительности жизни и среднего квадратичного отклонения. Таким образом. знание закона распределения длительности жизни обеспечивает наиболее компактную запись результатов эксперимента с минимальной потерей информации.
Следующая проблема, которая возникает перед экспериментатором, состоит в установлении достоверности различий выживаемости организмов при варьировании условий эксперимента. В современной геронтологии задача обычно "решается" путем механического использования методов параметрической статистики, (например, критерия Стьюдента),основанных на гипотезе о нормальном распределении продолжительности жизни. Между тем, как показали специальные исследования[Гаврилов,1980; Семенова, 1983], распределение организмов по срокам жизни резко отличается от нормального. Результаты этих исследований, основанные на сравнении наблюдаемых и теоретических распределений с помощью^-критерия, приведены в табл. 3.
Чтобы обеспечить высокую статистическую значимость результатов, были использованы таблицы смертности, построенные для популяций больших размеров (свыше 1000 особей). Можно заметить.
зз
Таблица 3
Несостоятельность нормального закона для описания распределения продолжительности жизни организмов
|
Характеристика генотипа
|
Исходная численность популяции
|
Х наблюдаемые значения
|
ХВД99 типичные критические значения
|
|
|
Drosophila melanogaster
|
|
|
|
Canton-S, самки
|
2400
|
1190,70
|
43,82
|
|
Canton-S, самки
|
1200
|
413,60
|
49,70
|
|
Canton-S, самцы
|
1200
|
190,96
|
48,27
|
|
Line 107, самки
|
1415
|
239,24
|
34,53
|
|
Line ЮТ, самцы
|
1407
|
212,37
|
32,91
|
|
vg^tt-pr/vj^b-pr, самцы
|
1061
|
190,75
|
26,13
|
|
Canton-S, самцы
|
2400
|
429,73
|
51,18
|
|
vg/vg, самцы
|
1048
|
165,24
|
26,13
|
|
purple-speck, самки
|
1750
|
107,37
|
39,25
|
|
vg/vg, самки
|
1026
|
174,37
|
22,46
|
|
purple, самки
|
1006
|
110,83
|
36,12
|
|
purple-speck, самцы
|
1465
|
135,41
|
40,79
|
|
vg^b-pr/vg^b-pr, самки
|
1099
|
198,48
|
27,88
|
|
Quintuple, самки
|
988
|
210,67
|
58,30
|
|
Quintuple, самцы
|
1000
|
216,20
|
59,71
|
|
Крысы линии Вистар
| |||
|
самцы
|
1430
|
116,24
|
37,70
|
|
самки
|
1045
|
114,69
|
40,80
|
что во всех случаях рассчитанные величины ^-критерия превышали соответствующие табличные критические значения. Это означает, что нормальный закон распределения продолжительности жизни с доверительной вероятностьюР >0,999 может быть отброшен как не соответствующий экспериментальным данным Отсюда следует, что выводы,полученные с использованием методов параметрической статистики, особенно в случае малых выборок, имеют лишь видимость научной строгости и должны быть пересмотрены заново. Таким образом, знание истинного закона распределения необходимо не только для того, чтобы установить достоверность различий при минимальном числе наблюдений, но и для того, чтобы избежать ошибочных выводов.
Наконец, на заключительном этапе исследований необходимо корректно интерпретировать полученные результаты. К сожалению, в этой области в настоящее время отсутствуют единые принципы интерпретации данных по выживаемости организмов. Так. нередко делается вывод о замедлении старения на основании данных об
34
увеличении продолжительности жизни организмов, а вещества, приводящие к такому увеличению, называют геропротекторами,т.е защищающими от старения (Обухова. Эмануэль, 1984] Между тем, как справедливо отмечается рядом авторов[Лэмб,1980], далеко не всякое воздействие, влияющее на смертность и продолжительность жизни, должно влиять на старение В противном случае в числогеропротекторовследовало бы записать пенициллин и всю массу социально-гигиенических мероприятий, которые привели к резкому увеличению длительности жизни людей в XX в Этот пример показывает. что в настоящее время необходима модель выживаемости организмов, которая была бы достаточно общей, чтобы быть признанной большинствомгеронтологов,но достаточно конкретной чтобы позволять проводить единообразную интерпретацию результатов эксперимента. Нетрудно заметить, что создание подобной модели предполагает знание хотя бы самых общих свойств распределения продолжительности жизни организмов