

Лекция 4.2 Изостерические и аллостерические механизмы регуляции активности ферментов
Регуляция активности ферментов в клетке, прежде всего, осуществляется внутриклеточными компонентами – субстратами, интермедиатами и продуктами различных метаболических путей.
Изостерическая регуляция
В роли изостерических регуляторов активности ферментов выступают субстраты, кофакторы, структурные аналоги и продукты реакции. Все перечисленные соединения являются соединениями, взаимодействующими с активными центрами ферментом, и поэтому их относят к изостерическим регуляторам.
Концентрация субстрата. Скорость любой энзиматической реакции определяется, в первую очередь, концентрацией субстрата. У ферментов, подчиняющихся закону Михаэлиса-Ментен, скорость реакции растет с увеличением концентрации субстрата, но достигает Vmax, когда фермент насыщен субстратом.
Vmax·[S]
v = --------------
Кm + [S]
Скорость энзиматической реакции наиболее чувствительна к изменению концентрации субстрата, когда она ниже Кm. При концентрации субстрата меньше, чем 1/10Кm, удвоение субстрата дает примерно удвоение скорости; при концентрации субстрата, равной 10Кm, удвоение концентрации субстрата оказывает малый эффект на скорость реакции. Если фермент имеет высокую Кm, то для достижения ½ Vmax ему требуются более высокие концентрации субстрата.
Например, гексокиназа I – изозим в эритроцитах имеет Кm для глюкозы ≈ 0,01 мМ, Кm для глюкозы глюкокиназы (изозим IV, печень) ≈ 10 мМ.
Эритроциты полностью зависят от метаболизма глюкозы, которую используют для образования АТР. При низких значениях Кm гексокиназы эритроцитов, даже если концентрация глюкозы в крови драматически снизится, эритроциты будут фосфорилировать глюкозу со скоростью, близкой к максимальной.
Печень же сохраняет излишек глюкозы в виде гликогена или превращает в жиры. Так как ее Кm ≈ 5 мМ, скорость фосфорилирования глюкозы будет увеличиваться с увеличением концентрации глюкозы в крови и уменьшаться, когда уровень глюкозы в крови падает. Высокая Кm гексокиназы печени поэтому способствует сохранению в печени гликогена или жиров, но только в случае избытка глюкозы.
Концентрация ингибиторов. Ферменты достаточно часто обратимо ингибируются структурными аналогами и продуктами реакции. Ингибиторы, похожие по структуре на субстрат конкурирует с ним за активный центр фермента, поэтому конкурентные ингибиторы увеличивают кажущуюся Кm фермента, но не влияют на Vmax. Они увеличивают концентрацию субстрата, необходимую для насыщения фермента, т.е. для достижения максимальной активности.
Классическим примером изостерического ингибирования является действие малоната на активность сукцинатдегидрогеназв (СДГ). Для 50 % ‒ го угнетения СДГ соотношение сукцинат/малонат должно составить 1:50. Изостерическим ингибитором СДГ является так же оксалоацетат, который регулирует, таким образом, заключительные этапы цикла Кребса по типу обратной связи.
Изостерические ингибиторы известны и для ферментов, метаболизирующих витамины. Примером может служить тиаминкиназа, фермент, осуществляющий фосфорилирование витамина В1, тиамина. При этом образуется активная форма тиамина ‒ тиаминдифосфат, участвующий в качестве простетической группы в функционировании пируватдегидрогеназы – первого фермента в составе пируватдегидрогеназного комплекса. Синтетический аналог тиамина, у которого атом серы заменен, на винильную группу – С=СН- , назван пиритиамином. Пиритиамин является одним из сильнейших изостерических ингибиторов. В присутствии тиамина пиритиамин ингибирует тиаминкиназу на 50% в соотношении 1:1. Следовательно, пиритиамин обладает таким же сродством к активному центру тиаминкиназы, как и тиамин, однако комплекс тиаминкиназа-пиритиамин катилитически не активен.
Все продукты являются обратимыми ингибиторами ферментов. Они могут быть конкурентными, неконкурентными, смешанными в отношении субстрата. Простейшее ингибирование продуктом, уменьшение скорости энзима, вызванное накоплением его собственного продукта, играет важную роль в метаболических путях. Это ингибирование не позволяет ферменту в последовательности реакций образовывать большее количество продукта, чем он может быть использован следующим ферментом в последовательности данного метаболического пути.
Регуляция активности аллостерических ферментов
Аллостерическая регуляция основана на изменениях конформации фермента, ведущей к изменению его активности. Аллостерическая регуляция является одним из самых быстродействующих и гибких типов модуляции активности ключевых метаболических ферментов. Она осуществляется с помощью молекул-эффекторов, взаимодействующих с аллостерическим центром фермента. Обычно аллостерический фермент построен из субъединиц, которые могут существовать в активной и неактивной конформации, и аллостерический эффектор промотирует превращение одной конформации в другую.
Аллостерическими эффекторами могут выступать самые различные вещества: субстраты и конечные продукты метаболических путей, иногда – промежуточные метаболиты; в катаболических процессах ‒ нуклеозиддифосфаты и нуклеозидтрифосфаты, а также переносчики восстановительных эквивалентов; в каскадных реакциях сАМР и сGMP, которые регулируют активность ферментов (например, протеинкиназ), участвующих в ковалентной модификации белков; ионы металлов и множество иных соединений.
Рассмотрим регуляцию аллостерических ферментов на примере фосфофруктокиназы и аспартаткартамоил-трансферазы (АКТФ).
Фосфофруктокиназа (ФФК) – один из наиболее хорошо изученных аллостерических ферментов. Это тетрамер, каждая субъединица которого содержит активный центр. Аллостерические центры располагаются на поверхности контакта двух субъединиц (рис. 4.2.1).

Рис. 4.2.1. Строение фосфофруктокиназы
Фосфофруктокиназа – поливалентный аллостерический фермент, активность которого регулируется разными внутриклеточными соединениями. Ингибиторами ФФК являются АТР, цитрат и концентрация протонов водорода. АТР является одновременно субстратом и аллостерическим ингибитором этого фермента.
Высокие уровни АТР изменяют кинетическое поведение ФФК, превращая гиперболическую зависимость (область низких концентраций АТР) в сигмоидную (рис. 4.2.2).

Рис. 4.2.2. Зависимость скорости фосфофруктокиназной реакции от концентрации фруктозо-6-фосфата в области низких и высоких концентраций АТР
Аллостерическими активаторами ФФК являются АМР и фруктозо-2,6-дифосфат, которые могут снимать ингибирующий эффект АТР. На рис. 4.2.3 показано влияние фруктозо-2.6-дифосфата на зависимость скорости реакции, катализируемой фосфофруктокиназой, от концентрации субстратов – фруктозо-6-фосфата и АТР.
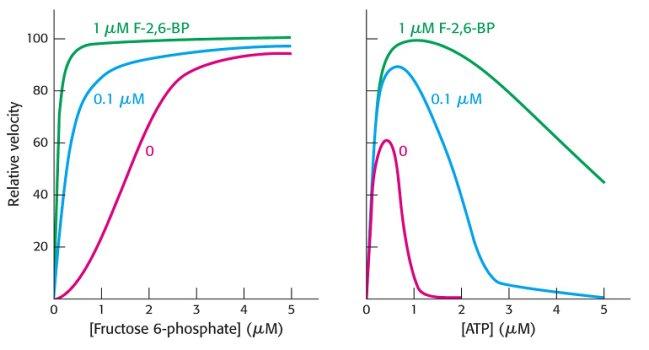
Рис. 4.2.3. Влияние различных концентраций фруктозо-2,6-дифосфата на зависимость скорости фосфофруктокиназной реакции от концентрации фуктозо-6-фосфата и АТР
Аспартаткарбамоилтрансфераза (АКТФ) – фермент, катализирующий скорость-лимитирующую реакцию в анаболическом пути синтеза пиримидиновых нуклеотидов. Она катализирует перенос карбамоильного остатка с карбамоилфосфата на аминогруппу L-аспартата. Образующийся N-карбамоил-L-аспартат содержит уже все атомы будущего пиримидинового кольца. Бактериальная АКТФ E. coli состоит из 12 субъединиц, шесть из которых являются каталитическими, другие шесть – регуляторными. На рис. 4.2.4 приведена субъединичная структура АКТФ (каталитические субъединицы окрашены в голубой и зеленый цвет, регуляторные – оттенки желтого).
Последние связывают аллостерические эффекторы CTP и АТР. Как и гемоглобин АКТФ может существовать в двух конформациях: менее активном Т-состоянии (от англ. tense — напряженное) и более активном R-состоянии (от англ. relaxed — расслабленное). Субстрат и эффекторы влияют на равновесие между обоими состояниями и тем самым на сигмоидность кривой. С возрастанием концентрации аспартата равновесие смещается к R-форме. АТР стабилизирует R-состояние путем связывания с регуляторной субъединицей. Напротив, присоединение СТР содействует переходу в Т-состояние. Структурные перестройки между R- и Т-состояниями особенно драматичны в случае АКТФ, поскольку при Т→R-переходе каталитические субъединицы удаляются друг от друга на 1,2 нм; кроме того, субъединицы поворачиваются вокруг оси симметрии. При этом сами конформации субъединиц меняются незначительно.

Рис. 4.2.4. Структура аспартаткарбамоилтрансферазы
Аллостерические эффекторы в зависимости от природы фермента могут влияять на максимальную скорость реакции – Vmax, на [S]0,5 и коэффициент Хилла – h. Если изменяется преимущественно Vmax, то фермент относится к так называемому V-типу ферментов. Ферменты, у которых у которых аллостерические эффекты отражаются только на [S] 0,5 и h, относятся к так называемому, К-типу аллостерических ферментов. АКТФ, наряду с гемоглобином, относится к К-типу аллостерических ферментов.
АКТФ имеют сигмоидную (S-образную) кривую насыщения субстратом – аспартатом. Активность фермента ингибируется цитидинтрифосфатом CTP — конечным продуктом анаболического пути обмена пиримидинов и активируется начальным участником — АТР. Ингибитор СТР вызывает смещение кривой зависимости скорости реакции от концентрации аспартата вправо с возрастанием [S]0,5 и h (рис. 4.2.5, кривая II). Активатор АТР, напротив, вызывает смещение влево; он уменьшает как [S] 0,5, так и h (рис. 4.2.5, кривая III).

Рис. 4.2.5. Зависимость скорости реакции, катализируемой аспартаткарбамоилтрансферазой (I – нативный фермент; II – фермент в присутствии СТР; III – фермент в присутствии АТР)
При регуляции мультиферментных комплексов таких, как, например, пируватдегидрогеназный комплекс, аллостерические механизмы осуществляются как участниками процесса окислительного декарбоксилирования пирувата (рис. 4.2.6 А), так и внешними по отношению к данному процессу, метаболитами (рис. 4.2.6 Б).

Рис. 4.2.6. Регуляция активности пируватдегидрогеназы (А – аллостерическая регуляция пируватдегидрогеназы; Б – аллостерическая регуляция киназы пируватдегидрогеназы; ковалентная модификация пируватдегидрогеназы ( + –положительный эффект, - отрицательный)
Регуляция метаболических путей
Большинство метаболических путей регулируется на уровне скорость-лимитирующих ферментов через различные механизмы, связанные с изменениями конформации фермента, действующими на каталитический участок. Эти регуляторные механизмы включают: 1)аллостерическую активацию, либо ингибирование; 2) ковалентную модификацию; 3) белок-белковые взаимодействия между регуляторными и каталитическими субъединицами или между двумя белками; 4) протеолитическое расщепление.
Эти типы регуляции могут быстро осуществлять переход фермента от неактивной к полностью активной конформации. В этой лекции мы остановимся на регуляции метаболических путей с помощью аллостерического механизма. Остальные из вышеперечисленных типов регуляции рассматриваются в лекции 4.3.
Регуляторные механизмы для скорость-лимитирующего фермента пути всегда отражают функцию метаболического пути в соответствующей ткани. Регуляция по типу обратной связи (feedback regulation) – конечный продукт пути прямо или опосредованно контролирует скорость своего собственного синтеза. Примером метаболического пути, в котором конечный продукт – пиримидиновый нуклеотид (СТР) регулирует свое собственное образование, является биосинтез пиримидинов, а аспартаткарбамоилтрансфераза является регуляторным ферментом этого пути.
Опережающая регуляция (feedforward regulation) – контроль скорости пути субстратом. Примером ее является процесс гликолиза, к котором фруктозо-1,6-дифосфат, образующийся в ходе ФФК реакции, является аллостерическим активатором пируваткиназа, способствуя более интенсивному превращению пирувата.
Регуляция по типу обратной связи характерна для анаболических процессов, тогда как опережающая регуляция характерна для катаболических процессов.
Метаболические пути также регулируются через компартментализацию ферментов.