

Мультиферментные комплексы
В их состав входят разные ферменты, катализирующие последовательные этапы превращения какого-либо субстрата. Особенность подобных комплексов – прочность ассоциации ферментов. Образование мультиферментных комплексов имеет определенный биологический смысл и ряд преимуществ.
Организация функционально связанных ферментов в комплексы концентрирует каталитическую активность, позволяет достичь высоких концентраций интермедиатов и перехода субстратов с одного активного центра на другой с минимизацией диффузионных эффектов. Кроме того, присутствие ферментов в комплексе делает возможным координацию аллостерического контроля нескольких активностей. Пируватдегидрогеназный комплекс являяется растворимым мультиэнзимным комплексом, который изучен в деталях.
Пируватдегидрогеназный комплекс
Этот мультиферментный комплекс, имеющий молекулярную массу 4500 кДа, состоит из трех типов ферментов. Первый из них (E1) ускоряет реакцию декарбоксилирования пировиноградной кислоты. В состав комплекса входит 12 димерных молекул этого фермента. Второй и третий ферменты, катализирующие окислительно-восстановительные процессы при окислении пировиноградной кислоты, сосредоточены внутри мультиэнзимного комплекса. Один из них (Е3) представлен шестью димерными молекулами, другой (Е2) ‒ 24 протомерами (рис. 4.1.4).

Рис.4.1.4. Трехмерная струкатура пируватдегидрогеназного комплекса
В результате слаженного во времени и пространстве действия всех трех видов входящих в его состав ферментов мультиэнзимный комплекс с огромной скоростью осуществляет превращение пировиноградной кислоты (4.1.5).
Сравнительно недавно выявлена еще одна своеобразная черта в строении ферментов: некоторые из них являются полифункциональными, т.е. обладают несколькими энзиматическими активностями, но всего лишь одной полипептидной цепью. Эта единая цепь при формировании третичной структуры образует несколько функционально и стерически обособленных глобулярных участков ‒ доменов, каждый из которых характеризуется своей каталитической активностью. Такие ферменты называют мультиферментными конъюгатами.

Рис. 4.1.5. Процесс декарбоксилирования пирувата пируватдегидрогеназным комплексом
Мультиферментные конъюгаты
Примерами полифункциональных конъюгатов являются комплекс синтазы жирных кислот (рис. 4.1.5) и КАД – комплекс, объединяющий первые три энзиматические активности. участвующие в метаболизме пиримидинов.
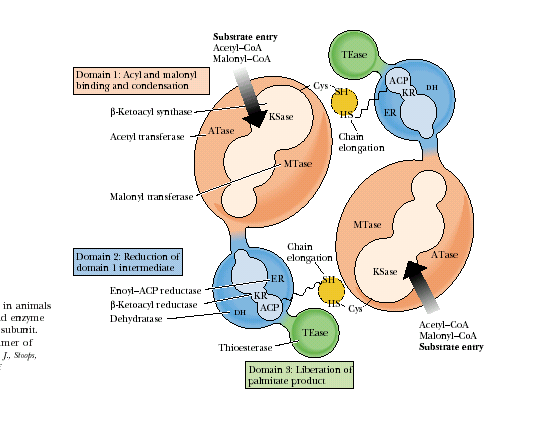
Рис.4.1.5. Комплекс синтазы жирных кислот млекопитающих
Каждая субъединица этого конъюгата включает три различных домена и восемь субдоменов.
Домен I состоит из трех субдоменов: субдомен 1 − АПБ-S-ацетилтрансфераза, 60 кДа; субдомен 2 − АПБ-S-малонилтрансфераза, 23 кДа; субдомен 3 − β-кетоацил-АПБ-синтаза (конденсирующий фермент, 45 кДа). Домен I катализирует присоединение субстратов ацетил-СоА и малонил-СоА ацетилтрансферазой и малонилтрансферазой соответственно и последующую конденсацию обоих партнеров β-кетоацил-синтазой.
Домен II также состоит из трех субдоменов: субдомен 4 − β-кетоацил-АПБ-редуктаза, 21 кДа; субдомен 5 − β-гидроксиацил-АПБ-дегидратаза, 50 кДа; субдомен 6 − еноил-АПБ-редуктаза, 14 кДа. К субдомену 4 присоединен ацилпереносящий белок (АПБ), 15 кДа. Домен II восстанавливает растущую цепь ЖК с помощью вышеназванных трех ферментов.
Домен III содержит субдомен 7 − ацил-АПБ-гидролаза, тиоэстераза, 33 кДа. Домен III после семь циклов удлинения цепи катализирует высвобождение готового продукта − пальмитата с помощью гидролитического фермента тиоэстеразы.
В настоящее время показано, что многие ферменты организованы в мультиферментные ансамбли. По мере роста наших знаний в деталях, число их увеличивается. Сплайсинг мРНК осуществляется в сплайсосомах, транскрипция осуществляется в транскрипционных комплексах. В синтез белка вовлечено множество ферментов, мРНК и рибосомы, образующие трансляционный комплекс. Деградация белков осуществляется протеосомами.
Кроме того, существуют доказательства мультиэнзимных систем в других путях (обозначенных метаболонами, от слова метаболизм ‒ обмен веществ), таких как гликолиз, цикл лимонной кислоты, синтез нуклеотидов, синтез мочевины, ЦТК, окисление жирных кислот и аминокислотный метаболизм.