

Смешанный тип ингибирования
Типы ингибирования, рассмотренные выше, являются предельными случаями в широком спектре возможных эффектов. В случае двусубстратных реакций, при определенной концентрации ингибитора, часто можно наблюдать ингибирование смешанного типа.
Общая схема ингибирования смешанного типа.

При этом Ki1 ≠ Ki2. Изменяются как константа Михаэлиса, так и максимальная скорость, но не в одинаковой степени:
![]() (8)
(8)
Смешанное конкурентное-неконкурентное ингибирование. В этом варианте ингибирования Ki2˃ Ki1. Сродство фермента к субстрату в присутствии ингибитора данного типа увеличивается, а максимальная скорость ферментативной реакции снижается (2.3.14).
 ,
,
 (9)
(9)
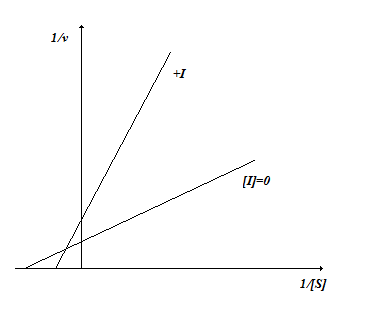
Рис. 2.3.14. Смешанное конкурентное-неконкурентное
Смешанное неконкурентное-бесконкурентное ингибирование. В этом случае Ki1 ˃ Ki2. Этот ингибитор снижает и константу Михаэлиса и максимальную скорость реакции (рис. 2.3.15).
 ,
,
 (10)
(10)

Рис. 2.3.15. Смешанное неконкурентное-бесконкуоентное.
Примером смешанного ингибирования является воздействие ртутьорганического соединения мертиолата на сахаразу грибов. Это вещество широко используется в промышленности для подавления роста микромицетов.
Субстратное ингибирование
Для многих ферментативных реакций при увеличении концентрации субстрата начальная скорость ферментативной реакции проходит через максимум, а затем уменьшается. Подобного рода зависимость можно описать, исходя из предположения об образовании в процессе реакции непродуктивного тройного комплекса ЕS2. В этом случае кинетическая схема ферментативной реакции выглядит следующим образом:

Зависимость скорости реакции от концентрации субстрата после математической обработки этой схемы имеет вид:
![]() (11)
(11)
Где К′s – субстратная константа, учитывающая сродство фермента ко второй молекуле субстрата. Анализ этого уравнения, как и уравнения Михаэлиса-Ментен, целесообразно проводить раздельно в области низких ([S] < Кs´) и высоких ([S] ˃ Кm ) концентрациях субстрата.
При низких значениях концентрации субстрата уравнение упрощается до классического уравнения Михаэлиса-Ментен (рис. 2.3.16а), при линеаризации которого можно определить Кm, Vmax, kcat (k+2 ).
![]()
В области высоких концентраций субстрата уравнение принимает вид:
![]() (12)
(12)
Линеаризация этого уравнения в координатах (1/v, [S]) позволяет определить Vmax и Кs´ (рис.2.3.16б).
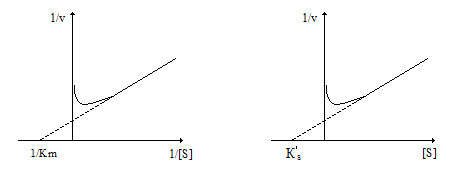
а) б)
Рис. 2.3.16. Линеаризация уравнения (12) в координатах (1/v 1/[S]) –а) и (1/v ; [S]) – б)
Зная величины Кm и Кs´, можно рассчитать оптимальную концентрацию субстрата по формуле:
![]() (13)
(13)
Методы определения константы ингибирования. Метод Диксона
Величины Кi можно измерить различными методами, как экспериментально, так и при помощи расчетов.
Если известна только одна концентрация ингибитора, то константу ингибирования можно определить, используя графики в координатах Лайнуивера-Берка. Найдя кажущиеся и истинные значения константы Михаэлиса и максимальной скорости для двух графиков (в отсутствии и в присутствии ингибитора) и подставив эти значения в формулы (2), (4), (7), (9), (10).
Наиболее удобен для определения константы ингибирования метод Диксона. Этот простой графический метод позволяет определять Кi непосредственно. Если определять скорость реакции в условиях постоянной концентрации субстрата и различных концентраций ингибитора, то график зависимости 1/v от [I] будет представлять собой прямую (рис. 2.3.17).

Рис. 2.3.17. Графическое определение ингибиторных констант (метод Диксона). Во всех случаях S1˃S2
В этом случае для определения ингибиторной константы достаточно определить скорость реакции всего для двух концентраций субстрата.