

40.Факторы регуляции численности популяции.
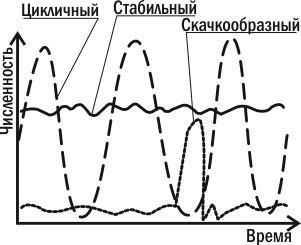
Три типа популяционной динамики.
Можно заключить, что численность каждой популяции находится под контролем множества отрицательных обратных связей. Когда численность популяции начинает расти, ее сдерживают и недостаток собственных ресурсов, и переключение на питание ее особями хищников-полифагов, и избыточное размножение специализированных хищников, и рост паразитарных инфекций. Все эти факторы «возвращают» численность популяции к прежнему уровню и снижают давление на нее, когда эта численность оказывается невысокой. В зависимости от того, насколько существенным окажется отклонение численности популяции от нормы, к ее возвращению подключится большее или меньшее количество регулирующих ее факторов
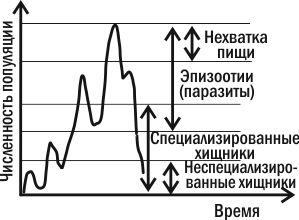
Из всех причин, вызывающих колебания численности, некоторые исследователи главную роль отводят климатическим условиям, считая биотические факторы второстепенными. По мнению других, только те факторы, которые зависят от плотности, вызывают колебания численности. Многие ученые признают роль тех и других.
В экосистемах с низким уровнем видового разнообразия численность популяций подвержена сильному воздействию физических стрессоров и зависит от погоды, химического состава среды и степени ее загрязнения. В экосистемах с высоким уровнем видового разнообразия колебания численности популяций в основном контролируются биотическими факторами.
Все экологические факторы в зависимости от характера их влияния на численность популяции можно разделить на две группы: 1) факторы, независимые от плотности; 2) факторы, зависимые от плотности.
К первой группе относятся факторы, действующие на популяции постоянно и изменяющие их численность однонаправленно, независимо от величины популяции. Так действуют абиотические факторы и прежде всего климатические факторы, а также антропогенные факторы, за исключением природоохранных факторов. Действие их не всегда проявляется сразу. Эти факторы еще называют модифицирующими, поскольку они не обеспечивают регуляции численности, а лишь вызывают ее изменения.
Ко второй группе относятся факторы, действие которых на популяцию может быть прямым, т.е. усиливаться по мере приближения к верхнему пределу, или инверсным, т.е. ослабевать по мере увеличения плотности. Эти факторы изменяют численность в сторону оптимального уровня и предотвращают перенаселение, поэтому В.Викторов предложил называть их регулирующими факторами. К ним относятся биотические факторы и природоохранные антропогенные факторы. Зависимые от плотности факторы, как правило, влияют на скорость роста численности популяции путем изменения ее рождаемости или смертности.
Биотические факторы, зависимые от плотности, по характеру влияния на численность популяции подразделяются на две группы:
1) внутривидовые;
2) межвидовые.
Рассмотрим характер действия внутривидовых факторов. К.Вини-Эдвардс в 1962 году предложил два механизма стабилизации плотности ниже уровня насыщения за счет внутривидовых факторов:
1) территориальное поведение;
2) групповое поведение.
При нарушении границ за счет увеличения плотности наблюдаются разные формы внутривидовой конкуренции:
1) конкуренция за ресурсы. При недостатке жизненно необходимых ресурсов часть животных погибает, а у растений наблюдается самоизреживание и изменение вегетативной мощности;
2) прямой антагонизм - биологическая и химическая «война» в природе. Биологическая «война» - это умерщвление конкурентов внутри популяции путем прямого нападения. Химическая «война» - это выделение химических веществ, задерживающих рост и развитие или убивающих молодых особей (растения, водные животные).
41.Регуляция численности на популяционном уровне. Синтетическая теория колебания численности.
новное и принципиально новое положение синтетической теории И.Я. Полякова состоит в том, что она позволяет заблаговременно судить о динамике численности и о вероятных факторах, способных на нее воздействовать, по состоянию кормовой базы, физической среды и морфофизиологическим свойствам популяций. Это делает ее приемлемой для решения задач прогноза численности вредителей (Поляков, 1976).
В последние годы 20-го столетия среди отечественных и зарубежных экологов были популярны теоретические концепции, названные Г.А. Викторовым (1973) стохастизмом и регуляционизмом, а современный этап исследований популяционной динамики поиском механизмов регуляции численности.
Представление о том, что каждая популяция обладает равновесным уровнем плотности и существуют выработавшиеся в ходе эволюции внутрипопуляционные (или внутриэкосистемные) механизмы, направленные на поддержание этой плотности, лежит в основе подхода, который наиболее точно можно охарактеризовать) как регуляционизм.
Для формирования данного подхода большое значение имели выполненные в 30—50-х гг. теоретические и экспериментальные работы австралийского энтомолога А. Никольсона (Nicholson, 1933, 1957), подчеркнувшего, что динамика численности любой популяции есть автоматически регулируемый процесс, а действие, факторов, контролирующих популяцию, управляется плотностью самой контролируемой популяции. Выше уже говорилось об опытах Никольсона с падальной мухой, наглядно продемонстрировавших механизм возникновения в популяциях автоколебаний численности.
Принципиально другой подход — это стохастизм, уделяющий основное внимание факторам, случайно действующим, или, точнее, случайно распределенным во времени и в пространстве. Сторонники стохастизма обычно отрицают существование «равновесного» уровня, всякое отклонение от которого будто бы автоматически включает процессы, возвращающие популяцию к исходному уровню. С позиции стохастизма «равновесный уровень численности (т. е. тот, при котором R0 = 1) есть просто артефакт усреднения за длительный срок: чем длиннее имеющийся ряд наблюдений за какой-нибудь популяцией, тем больше шансов утверждать, что средняя плотность, полученная за ряд лет, это и есть «равновесная» плотность, активно поддерживаемая специальными механизмами.
Спор сторонников регуляционизма и стохастизма достиг своей кульминации в конце 50-х гг., особенно после выхода в свет капитальной сводки «Распространение и обилие животных», принадлежащей перу австралийских исследователей Г. Андреварты и Л. Берча (Andrewartha, Birch, 1954). Проанализировав очень большой эмпирический материал (главным образом по насекомым), Андреварта и Берч пришли к выводу о том, что популяции животных в природе обычно ограничены: 1) нехваткой ресурсов (пищи, подходящих мест для гнездовий и т. д.); 2) недоступностью этих ресурсов вследствие ограниченных возможностей расселения животных; 3) кратковременностью периода, в течение которого скорость роста популяции r сохраняет положительное значение. По мнению сторонников стохастизма, распространение какого-либо вида организмов в пространстве и динамика численности его популяций во времени ограничены одними и теми же факторами, тогда как сторонники регуляционизма полагают, что распространение организмов лимитируется главным образом абиотическими (независимыми от плотности) факторами, а динамика численности — биотическими, как правило, зависимыми от плотности.
Соронники первого направления (Thompson, 1956; Bodenheimer, 1958; Andrewartha, Brich, 1954; Milne, 1984 и др.) считают воздействия факторов внешней среды на популяции случайными. Комбинации различных факторов определяют изменения численности насекомых (подъемы и спады), причем благоприятное сочетание условий, определяющих подъемы численности, наблюдается в природе гораздо реже, чем неблагоприятное.
Представители второго направления рассматривают колебания численности как регулируемый процесс. Они считают, что случайные изменения ее, вызванные прямым или косвенным воздействием абиотических (главным образом физических) факторов, компенсируются деятельностью регуляторных механизмов, которые управляются изменениями плотности популяций по принципу отрицательной обратной связи. По мнению сторонников регуляционизма, в этой роли могут выступать биотические факторы среды, реагирующие на изменения численности других организмов (Howard, Ficke, 1911; Varley, 1953; Lack, 1956; Winne-Edmards, 1965; Klomp, 1964; Уатт, 1971; Дажо, 1975; Ю. Одум, 1986; Уильямсон, 1975; Варли, Градуэлл, Хассел, 1978; Риклефс, 1979; Пианка, 1981).
Согласно представлениям большинства современных экологов, изменение численности насекомых рассматривается как взаимодействие различных механизмов. Г.А. Викторов (1967) подразделил их на модифицирующие и регулирующие. К модифицирующим он отнес климатические и другие географические факторы среды, к регулирующим — естественных врагов (паразиты, хищники, возбудители болезней), внутривидовые отношения (конкуренция), а также трофические факторы (количество, качество и доступность пищи).
Синтетическая теория в варианте И.Я. Полякова при всех ее положительных сторонах не решает проблему многолетних флюктуаций численности животных. Характерно, что она используется в практике защиты растений лишь для краткосрочных биологических прогнозов.
42-46.Классификация межвидовых взаимоотношений. Отрицательные и положительные взаимоотношения./Конкуренция и хищничество./Паразитизм и аллелопатия./Комменсализм, протокооперация, мутуализм./Консорции.
Возможны следующие виды влияний одних организмов на другие:
-
Положительное (+) — один организм получает пользу за счёт другого.
-
Отрицательное (−) — организму причиняется вред из-за другого.
-
Нейтральное (0) — другой никак не влияет на организм.
Симбиоз:
+ + Облигатный симбиоз — форма симбиоза, при которой в естественных условиях популяции не могут существовать друг без друга (пример: симбиоз гриба и водоросли в лишайнике).Разновидностью облигатного симбиоза является:
Мутуализм — форма облигатного взаимовыгодного сожительства организмов двух и более видов.
Примером мутуализма является симбиоз рыб-клоунов с актиниями. Вначале рыба слегка касается актинии, позволяя ей ужалить себя и выясняя точный состав слизи, которым покрыта актиния, — эта слизь нужна актинии, чтобы она сама себя не жалила. Затем рыба-клоун воспроизводит этот состав и после этого может прятаться от врагов среди щупалец актинии. Рыба-клоун заботится об актинии — вентилирует воду и уносит непереваренные остатки пищи. Рыбки никогда не удаляются далеко от «своей» актинии. Самцы прогоняют от неё самцов, самки — самок. Территориальное поведение, видимо, стало причиной контрастной окраски.
Самая тесная форма мутуализма — когда один организм живёт внутри другого. Поразительным примером этого служит система органов пищеварения коров и других жвачных животных. Коровы, как и человек, не способны переварить целлюлозу — вещество, которое в большом количестве содержится в растениях. Но у жвачных животных есть особый орган — рубец. Он представляет собой полость, в которой живут множество микробов. Растительная пища, после того как животное её прожевало, попадает в рубец, и там эти микробы разрушают целлюлозу. (Животное может отрыгнуть и вновь прожевать частично расщеплённую пищу — именно этим и занимаются коровы, когда пережёвывают свою жвачку.) Рубец коровы — это замкнутая микроэкосистема, образованная множеством различных микроорганизмов, задача которых состоит в переработке целлюлозы для своего хозяина. Аналогично корневая система высших растений образована переплетением корневой ткани и грибных нитей, так что грибы снабжают растение минеральными веществами.
Мутуализм может быть «жёстким» или «мягким». В первом случае сотрудничество жизненно необходимо для обоих партнёров (они связаны отношениями коадаптации), во втором отношения более или менее факультативны (это называется протокооперацией).
+ + Факультативный симбиоз (Протокооперация (англ.)) — форма симбиоза, при которой совместное существование выгодно, но не обязательно для сожителей. (например, взаимоотношения краба и актинии: актиния защищает краба и использует его в качестве средства передвижения)
0 + Комменсализм — форма симбиоза, при которой одна популяция извлекает пользу от взаимоотношения, а другая не получает ни пользы, ни вреда. Выделяют следующие типы комменсализма: зоохорию, паройкию, синойкию (квартирантство), энтойкию, эпибиоз, эпиойкию (эпойкию, нахлебничество) , форезию.
Комменсализм (от лат. com — «с», «вместе» и mensa — «стол», «трапеза»; буквально «у стола», «за одним столом»; ранее — сотрапезничество) — способ совместного существования (симбиоза) двух разных видов живых организмов, при котором один из партнёров этой системы (комменсал) возлагает на другого (хозяина) регуляцию своих отношений с внешней средой, но не вступает с ним в тесные взаимоотношения[1]. При этом, популяция комменсалов извлекает пользу от взаимоотношения, а популяция хозяев не получает ни пользы, ни вреда (например, чешуйница обыкновенная и человек), т.е. метаболические взаимодействия и антагонизм между такими партнёрами чаще всего отсутствуют. Комменсализм — как бы переходная форма от нейтрализма к мутуализму.
Классификация:
-
паройкия — один организм использует другого (его самого, либо его жилище: раковину, гнездо и т. п.) в качестве убежища; складывается между животными, обладающими средствами защиты и незащищенными (например, некоторые окунеобразные прячутся между щупальцами крупных актиний, а те питаются остатками пищи рыб);
-
синойкия (квартирантство) — один организм (комменсал) использует другого (его самого либо его жилище) в качестве жилища;
-
энтойкия — одни животные поселяются внутри полостей других, имеющих сообщение с внешней средой, например рыбки Carapus обитают в клоаке голотурий, но питаются во внешней среде[3].
-
эпибиоз — одни организмы живут на поверхности других (например, усоногие рачки на горбатых китах, непаразитические эпифиты на других растениях).
-
эпиойкия (эпойкия, нахлебничество) — один организм (комменсал) прикрепляется к организму другого вида или живёт возле него, используя остатки пищи хозяина (например, рыба-прилипала плавником-присоской прикрепляется к коже акул и других крупных рыб, передвигаясь с их помощью и питаясь остатками их трапезы), водоросли, живущие в шерсти ленивца. Эпиойкия является одним из путей перехода к паразитизму[4];
-
инквилинизм — одно животное (инквилин), проникая в чужое жилище, уничтожает его хозяина, после чего использует жилище в своих целях. Тесно связан с синойкией. Изначально инквилинизм выделялся как подвид комменсализма, хотя он очень близок к хищничеству и паразитизму;
-
зоохория — распространение диаспор при помощи животных;
-
форезия — расселение организма при помощи его переноса другим, (например, мелкие клещи нескольких семейств, личинки жуков Cryptophagidae, Meloidae, Rhipiphoridae). Важный признак истинной форезии — отсутствие между форонтом и транспортным хозяином физиологической или биохимической зависимости
− + Паразитизм — форма симбиоза, при которой один организм (паразит) использует другой (хозяин) в качестве источника питания или/и среды обитания, возлагая при этом (частично или полностью) на хозяина регуляцию своих отношений с внешней средой. Паразитизм так же бывает облигатным, когда паразит не может существовать без хозяина (типичный пример — вирусы) и факультативным (вши, блохи, паразитические черви и т. д.).
-
Инквилинизм — одно животное, проникая в чужое жилище, уничтожает хозяина, после чего использует жилище в своих целях.
Хищничество:
− + Хищничество — явление, при котором один организм питается органами и тканями другого (при этом умерщвление жертвы не обязательно), при этом не наблюдается симбиотических отношений. Но в современной экологии часто используется общее понятие хищничества, в которое также входят паразитизм и растительноядность (фитофагия).
Нейтрализм: