

3. Метод неселективного изъятия
Данный метод используется для оценки абсолютной численности животных, обитающих на ограниченной территории. Он может быть применен для оценки численности насекомых на определенном участке луга, млекопитающих в локальной популяции и т.п. В основе использования метода неселективного изъятия лежит явление постепенного снижения вероятности встречаемости животного в серии последовательных отловов, вызванное снижением численности популяции в результате изъятия из нее особей.
Таким образом, при применении данного метода животных отлавливают, подсчитывают их количество и не выпускают до конца исследования. Затем производят еще 3-4 последовательных отлова по аналогичной методике, при этом число отловленных животных постепенно уменьшается, вследствие уменьшения их общего количества на исследуемой территории. Если теперь построить график зависимости числа отловленных животных при каждом отлове от общего числа ранее отловленных, то по нему можно найти оценку исходной численности популяции (Грин и др., 1990).
Однако довольно часто встречаются случаи, когда особи обладают высокой миграционной активностью и место отловленных резидентных особей сразу же занимают иммигранты. (Подобная ситуация может встретиться, например, при отловах мелких млекопитающих.) Тогда совокупное число отловленных животных не будет снижаться при каждом последующем отлове, что даст заниженную оценку коэффициента b и, следовательно, завысит оценку начальной численности популяции.
Метод трехкратных отловов Бейли позволяет, помимо оценки численности популяции, оценить также показатели рождаемости и смертности в течение периода исследования (Bailey, 1951; 1952; Коли, 1979). Кроме того, он применим в том случае, если животные довольно активно перемещаются.
Для того чтобы оценить численность популяции с помощью метода Бейли необходимы иметь данные по двум последовательным мечениям и двум последовательным отловам. Таким образом, в момент t0 производится отлов выборки животных. Их всех единообразно метят и выпускают обратно. Спустя некоторое время в момент t1 отлавливается новая выборка, в ней подсчитывается количество меченых особей. Остальные (немеченые) животные из этой выборки также метятся, но на этот раз новой меткой, отличной от первой. Спустя время в момент t2 вновь отлавливается выборка животных и в ней подсчитывается число особей, меченых в первый раз и во второй.
Оценить численность популяции можно и проводя многократные отловы, при этом производя единообразное мечения всех ранее немеченых животных при каждом последовательном отлове (Poole, 1974). В этом случае можно ожидать, что с каждым последовательным отловом и мечением в популяции будет расти доля меченых особей вплоть до того момента, когда все особи в данной популяции окажутся с метками. Таким образом, на основании оценки доли меченых особей в популяции (полученной с помощью выборочных данных) в зависимости от совокупного числа всех ранее меченых животных можно найти оценку величины популяции – ее численность, при которой доля меченых животных должна достичь 100 %.
Метод Джолли-Зебера
Если при каждом последовательном отлове особям присваиваются индивидуальные метки, то в этом случае можно оценить численность популяции (а также целый набор других экологических популяционных показателей) на основе метода Джолли-Зебера (Jolly, 1965; 1982; Seber, 1965; Begon, 1979; Коли, 1979).
Использование этого метода требует, чтобы в распоряжении исследователя были подробные сведения относительно дат мечения и отловов каждого конкретного животного.
31-12.Рост и скорость роста популяции./ Рождаемость и смертность в популяции.
Естественные популяции — это не раз и навсегда застывшая совокупность особей, а динамическое единство находящихся во взаимоотношениях организмов. Изменение в численности, структуре и распределении популяций как реакция на условия окружающей среды называется динамикой популяции.
Динамика популяций в упрощенном варианте может быть описана такими показателями, как рождаемость и смертность. Это наиболее важные популяционные характеристики, на основании анализа которых можно судить об устойчивости и перспективном развитии популяции.
Рождаемость определяется как число особей, рожденных в популяции за некоторый промежуток времени (час, день, месяц, год). Термин «рождаемость» относится к особям любых видов, независимо от способов появления их на свет: будь это прорастание семян подорожника или овса, появление детенышей из яиц у курицы или черепахи, рождение потомства у слона, кита либо человека.
Экологи выделяют максимальную рождаемость в условиях отсутствия лимитирующих экологических факторов (добиться этого весьма сложно, даже невозможно). Под максимальной рождаемостью понимается теоретически возможный максимум скорости образования новых особей в идеальных условиях. Размножение организмов сдерживается только их физиологическими особенностями. Теоретическая скорость размножения различных видов может быть довольно высокой.
В отличие от максимальной экологическая, или реализованная, рождаемость (или просто рождаемость) характеризует прирост или увеличение численности популяции при фактических или специфических условиях среды.
Смертность — это число особей, погибших в популяции за единицу времени. Подобно рождаемости, смертность можно выразить числом особей, погибших за данный период (число смертей в единицу времени), или в виде удельной смертности для всей популяции (или ее части). При определении смертности популяции учитываются все погибшие особи независимо от причины смерти (умерли ли они от старости или погибли в когтях хищника, отравились ядохимикатами или замерзли и т.д.).
Общие изменения численности популяции складываются за счет четырех явлений: рождаемости, смертности, вселения и выселения особей (иммиграция и эмиграция).
рвая характеризуется общим числом родившихся особей. Удельную рождаемость вычисляют как среднее изменение численности особей за определенный период времени, деленное на их первоначальное число.
Величина рождаемости зависит от многих причин. Большую роль играет плодовитость особей. Смертность в популяциях также зависит от многих причин: генетически запрограммированной длительности жизни особей, их генетической и физиологической полноценности, влияния неблагоприятных физических условий среды, воздействия хищников, паразитов, болезней и т.п.
Наиболее часто в природе встречается повышенная смертность особей в ранний период жизни.
Расселение (выселение) особей из популяции или пополнение ее пришельцами — закономерное явление, основанное на одной из важнейших биологических черт вида — его расселительной способности. В каждой популяции часть особей регулярно покидает ее (дисперсия популяции), пополняя соседние или заселяя новые, еще не занятые видом территории.
Современная теория рассматривает темп роста численности популяций как авторегулируемый процесс. Любой популяции организмов в конкретных условиях свойствен определенный средний уровень численности, вокруг которого происходят колебания.
В одном случае темп роста с самого начала высокий и постоянный, не зависящий от нарастающей плотности, что соответствует лавинообразному, по экспоненте, увеличению численности популяции (рис. 6.2а). Его графически описывает простейшая кривая, характеризующая изменение численности популяции, движущейся к равновесию, при условии изобилии пищи. При достижении же определенной плотности рост популяции прекращается. Если лимитирующий фактор среды обитания действует весьма быстро, то рост популяции прекращается внезапно (кривая «Б» на рис. 626).
Однако среда обитания оказывает ограничивающее влияние не резко, а постепенно, что приводит к 5-образной кривой роста (рис. 6.2#). Такая форма кривой роста наблюдается при внедрении популяции в новую территорию. В этом случае вначале происходит ускоренный рост (по логарифмическому закону). Затем, под влиянием сопротивления среды обитания, рост замедляется, и в популяции наступает фаза равновесия.
Если же популяция испытывает внешние воздействия (например, нападение хищников), то при постоянной удельной скорости изъятия особей в природе устойчиво существуют взаимодействующие популяции хищник — жертва (кривая «Г» на рис. 6.2и), но при меньшем уровне численности: Nиз меньше Nmax.
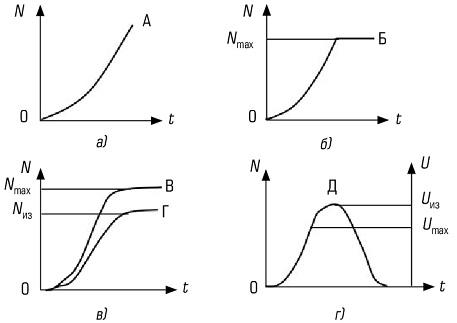
Кривые роста популяции: А — экспоненциальная; 5 — экспоненциальная с прекращением роста; В — логистическая; Г — логистическая с изъятием особей без превышения квоты; Д — с превышением квоты. N — численность популяции (Nmах — максимальная); (Uиз- реальная скорость изъятия, Umax — критическая для популяции скорость изъятия продукции; t — время
Удельная скорость изъятия — число изъятых особей в единицу времен, отнесенных к численности популяции. Если же человек изымает биопродукцию из популяции с постоянной (интегральной, но не удельной) скоростью, то возникает понятие квоты.
Следовательно квота представляет собой скорость отлова. Когда квота не превышает установленной критической величины, то равновесие популяции сохраняется. В этом случае отлов можно вести сколь угодно долго без губительных последствий для популяции. S-образная кривая называется логистической кривой роста, поскольку она получена путем интегрирования уравнения, основанного на логически обоснованных допущениях. Если квота превышает критическую величину отлова, то происходит полный отлов популяции за конечное время: популяция не успевает самовосстановиться и гибнет (рис. 6.2г)
Весьма интересны для ученых-экологов циклические популяции, подверженные закономерным колебаниям численности. Однако единой теории удовлетворительно объясняющей закономерности в циклических популяциях пока нет.
Поддержание определенной плотности получило название гомеостаза популяций. В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой она принадлежит. Механизмы популяционного гомеостаза зависят от экологической специфики вида, его подвижности, степени воздействия хищников и паразитов и др. У одних видов они могут проявляться в жесткой форме, приводя к гибели избытка особей, у других — в смягченной, например, в понижении плодовитости на основе условных рефлексов.
Другой механизм ограничения численности популяций — такие изменения физиологии и поведения при увеличении плотности, которые, в конечном счете, приводят к проявлению инстинктов массовой миграции.
Наиболее эффективным механизмом сдерживания роста численности популяции на данном ареале является определенная система инстинктов — мечение и охрана участков, не допускающие размножения на них «чужих» особей.
Генетические процессы в популяциях. В настоящее время известно, что все природные популяции гетерогенны и насыщены мутациями. Генетическая гетерогенность любой популяции при отсутствии давления внешних факторов должна быть неизменной, находиться в определенном равновесии.
Положение о генетическом единстве популяции является одним из наиболее важных выводов популяционной генетики: любая популяция представляет сложную генетическую систему, находящуюся в динамическом равновесии.
Прирост популяции - разница между рождаемостью и смертностью; прирост может быть положительным, нулевым и отрицательным.
Скорость роста популяции — изменение численности популяции в единицу времени. Зависит от показателей рождаемости, смертности, миграции и эмиграции.