

Подготовка у универсиаде 2012 / Генетика (Жимулев) / 15ver7
.pdfГенетика развития |
Глава 15 |
|||
|
|
|
||
Глава 15. Генетика развития |
|
|
||
|
Рисунок 15.1 |
|||
15.1. Преформизм |
|
|
||
и эпигенетика |
|
|
||
Вопрос о том, как из оплодотворенного |
|
|
||
яйцавырастаетцелыйорганизмикаквозникают |
|
|
||
различиямеждусоставляющимиегоклетками, |
|
|
||
интересует людей почти 2000 лет, но, тем не |
|
|
||
менее,досихпоростаетсяоднойизосновных |
|
|
||
проблем биологии и генетики развития в |
|
|
||
частности. |
|
|
|
|
 õîäå |
развития формируются |
|
|
|
многочисленныеорганыиткани,совершенноне |
|
|
||
похожиедругнадруга.Ониорганизованыдля |
|
|
||
выполненияопределенныхфункцийикаждая |
|
|
||
тканьпоразительноотличаетсяотостальных. |
|
|
||
Необходимо решить две проблемы: каким |
|
|
||
образом ткани дифференцируются и каким |
|
Миниатюрный человечек, формирующийся |
||
образом дифференцированное состояние, |
|
по мнению ранних биологов в сперматозоиде |
||
характерноедлякаждойклетки,наследуетсяв |
|
человека (Из: Hartsoeker, 1694, в кн. Srb et al., |
||
рядуклеточныхпоколений. |
|
1965, ñòð. 353). |
||
Долгое время в биологической науке |
определяютсяпротоплазмойяйца,влияниеже |
|||
существовало мнение, что процесс развития - |
хромосом сперматозоида сказывается позже. |
|||
это простой рост органов организма, уже |
Это означает, что протоплазма яйца уже |
|||
сформированного (преформированного) в |
подверглась влиянию генов, содержащихся в |
|||
клеткахзародышевогопути(Рис.15.1). |
самом яйце...” (Морган, 1937, стр. 129). “По |
|||
В 1759 году Wolff предложил теорию |
мерепротеканияразвитияприходятвдействие |
|||
эпигенеза,согласнокоторойкаждыйорганизм |
различныегруппыгенов”(тамже,стр.13). |
|||
развивается в ходе онтогенеза не из |
|
|
||
преформированных органов, а из простого |
Литература к разделу 15.1. |
|||
неорганизованного зародыша пут¸м |
АлиханянС.И.,АкифьевА.П.,ЧернинЛ.С.Общая |
|||
последовательногорядановообразований. |
|
генетика. Москва, Высшая школа, 1-446, |
||
|
|
|
||
По современным представлениям, |
1985. |
|||
жизненный путь любого организма - это |
Корочкин Л.И. Взаимодействие генов в развитии. |
|||
постоянное обновление всех клеток, тканей и |
Москва, Наука, 200 с., 1977. |
|||
органов. Детали этого процесса обновления |
Корочкин Л.И. Как гены контролируют развитие |
|||
определяются |
структурами, |
|
клеток. Соросовский образовательный |
|
сформировавшимисянапредыдущихстадиях. |
журнал 1: 17-22, 1996. |
|||
Согласно этой точке зрения, развитие не |
Корочкин Л.И. Введение в генетику развития. |
|||
останавливается на какой-то определ¸нной |
Москва, Наука, 1-252, 1999. |
|||
точке,апродолжаетсявсюжизнь. |
Лобашев М.Е. Генетика (издание второе). |
|||
Зависимостьразвитияотактивностигенов, |
|
Ленинград, Издательство ЛГУ, 1-751, 1967. |
||
Маркерт К., Уршпрунг Г. Генетика развития. |
||||
заключенныхвклеточномядре,установленав |
||||
многочисленныхопытах.Показанатакжероль |
Москва, Мир, 1-270, 1973. |
|||
Морган Т.Г. Развитие и наследственность. |
||||
состояния цитоплазмы в поддержании |
||||
определенного |
дифференцированного |
Ленинград, Биомедгиз, 1-241, 1937. |
||
Srb A.M., Owen R.D., Edgar R.S. General genetics. |
||||
состоянияклеткивцелом. |
|
Second edition. San Francisco, London, W.H. |
||
|
|
|
||
ПомнениюТ.Х.Моргана,высказанному
еще в 1930-е годы, “ранние стадии развития
Freeman and company, 1-577, 1965.
357
Глава 15 |
Генетика развития |
|
|
15.2. Роль клеточного ядра в развитии
Для выяснения роли ядра в развитии проводили различные эксперименты. Г. Геммерлинг(H.Haemmerling)провелопытыс замещениемядрауводорослиAcetabularia.Он использовал два вида этого рода - A. mediterranea и A. crenulata, различающиеся формой шляпки (Рис. 15.2). В период вегетативногоциклаэтаводоросльпредставляет собойкрупнуюодноядернуюклеткусдлиной стебелькадо6см.Различныевидыацетабулярии имеютспецифическуюформушапочки.
Ядронаходитсяводномизризоидов.Если шапочкуилистебелексшапочкойотрезать,они вновьрегенерируютизризоида,содержащего ядро. При этом сохраняется форма шапочки, характернаядляданноговида.Когдасращивали отрезанный стебелек одного вида с ризоидом другого,регенерирующаянастебелькешапочка имела форму, свойственную виду, которому принадлежало ядро (см. Рис. 15.2). Такой же результат был получен в том случае, когда извлеченное из ризоида ядро одного вида пересаживали в изолированный стебелек другоговида.
Б.Л. Астауров, основываясь на резко различнойчувствительностиядраицитоплазмы к ионизирующим излучениям, показал решающуюрольядравопределениипризнаков
Рисунок 15.2
Регенерация шляпки при перекрестном сращивании у ацетабулярии (Из: Hammerling, 1943 в кн. Srb et al., 1965). A. mediterranea и A. crenulata изображены соответственно серым и белым цветом.
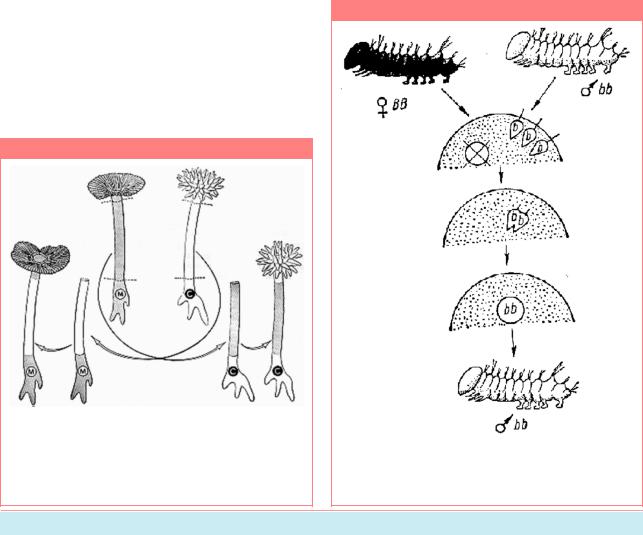
многоклеточныхорганизмов.Внормеубабочки шелкопряда в яйцо при оплодотворении проникаетнесколькоспермиев,носядромяйца сливаетсяядротолькоодногоизних,остальные спермии остаются на его периферии и затем распадаются, не участвуя в образовании и развитии зародыша. Подвергая неоплодотворенные яйца шелкопряда тепловомушокуирентгеновскомуоблучению, можно полностью разрушить их ядра, не повредивцитоплазму,котораяпосравнениюс ядром гораздо менее чувствительна к этим воздействиям.Еслизатемтакиеядраосеменить, то ядра двух проникших в ядро спермиев сливаются друг с другом и образуют ядро зиготы. Следовательно, здесь зигота имеет ядро, происходящее исключительно от отца, цитоплазма же целиком материнская. Развивающиеся из таких андрогенетических зиготособишелкопрядавсегдабылисамцами и довольно точно повторяют фенотип своих отцов, что особенно заметно, когда яйца принадлежатсамкамоднойпороды,аспермиисамцамдругой(Рис.15.3).
Рисунок 15.3
Андрогенез у тутового шелкопряда (По данным Б.Л. Астаурова в кн. Гершензон, 1983, стр. 83).
358

Генетика развития |
|
|
|
Глава 15 |
|
|
|
|
|
Литература к разделу 15.2. |
|
|
|
|
|
Рисунок 15.3 |
|
|
|
Астауров Б.Л. Партеногенез, андрогенез и |
|
|
|
|
полиплоидия. Москва, Наука, 1-344, 1977. |
|
|
|
|
Гершензон С.М. Основы современной генетики. |
|
|
|
|
Киев, Наукова Думка, 1-558, 1983. |
|
|
|
|
Корочкин Л.И. Введение в генетику развития. |
|
|
|
|
Москва, Наука, 24-40, 1999. |
|
|
|
|
Srb A.M., Owen R.D., Edgar R.S. General genetics. |
|
|
|
|
Second edition. San Francisco, London, W.H. |
|
|
|
|
Freeman and company, 1-577, 1965. |
|
|
|
|
15.3. Тотипотентность |
|
|
|
|
генома |
|
|
|
|
До сих пор обсуждается вопрос о том, |
|
|
|
|
сопровождается ли специализация клеток |
|
|
|
|
животных утратой генов, которые далее не |
|
|
|
|
понадобятся для каждого данного типа |
|
|
|
|
клеток. Например, сохраняются ли в ядре |
|
|
|
|
клеток кишечника гены, необходимые для |
|
|
|
|
синтеза гемоглобина - белка, характерного |
|
|
Äæîí øðäîí |
|
для эритроцитов, а в ядре нервной клетки - |
|
|
||
|
|
ðîä. 1933 |
|
|
|
|
|
|
|
гены,необходимыедляобразованиямиозина |
|
|
|
|
- особого белка мышечных клеток? Если |
двух ядрышек в ядрах клетки-хозяина и |
|||
“ненужные” гены утрачиваются, то именно |
одного - в пересаживаемых ядрах. |
|||
эта постоянная утрата различных генов и |
|
Вданныхэкспериментахоколо 1% яиц, |
||
определяет специализацию клеток, как |
в которые были пересажены ядра эпителия, |
|||
предположил А. Вейсман еще в 1892 году. |
дали взрослых лягушек. Таким образом, |
|||
Противоположная точка зрения сводится к |
нормальная дифференцировка клеток у |
|||
тому, что во всех клетках сохраняются все |
животных не сопровождается утратой или |
|||
гены, однако в тех клетках, где их |
необратимойинактивациейгенов. |
|||
деятельность не нужна, они находятся в |
|
В феврале 1997 года в журнале Nature |
||
неактивном состоянии. |
появилось |
сообщение, заставившее |
||
Для того, чтобы решить, какая из этих |
обсуждать проблемы генетики развития |
|||
гипотез верна, английский генетик Дж. |
журналистов, политиков, юристов и |
|||
Г¸рдон (Рис. 15.4) трансплантировал ядра из |
государственныхдеятелей:группауч¸ныхиз |
|||
специализированных клеток эпителия |
Шотландии |
сообщила |
об успешной |
|
кишечника головастиков шпорцевой |
трансплантацииядериздифференцированных |
|||
лягушки Xenopus laevis (Рис. 15.5) в |
клеток в яйцеклетки овец и получении |
|||
неоплодотвор¸нное яйцо этого же вида; из |
нормальносформированногоживотного.Эти |
|||
яйца которого предварительно было удалено |
результаты открывают путь для фактически |
|||
свое ядро. При этом необходимо |
неограниченного |
вегетативного |
||
использование какого-либо ядерного |
размножения любого индивидуума: каждая |
|||
маркера, который позволяет отличить |
особьврезультатетрансплантацииядеризего |
|||
потомков пересаженного ядра от потомков |
клеток в реципиентов может дать начало |
|||
ядра-хозяина, поскольку нельзя быть |
миллиардам совершенно идентичных |
|||
уверенным, что не произошло случайной |
потомков. Этот процесс, как отмечалось в |
|||
ошибки при удалении ядра-хозяина. |
разделе 4, называют клонированием. |
|||
Маркером в данном случае служило наличие |
|
|
|
|
359

Глава 15 |
|
Генетика развития |
|
Рисунок 15.5 |
|
|
|
1 |
2 |
Головастик |
|
(линия с одним ядрышком) |
|||
|
|
Неоплодотворенное яйцо (линия с двумя ядрышками) Кишечник
 головастика
головастика
Облучение |
Эпителиальные клетки |
|
|
||
|
3 |
Ядро из клетки |
|
|
|
4 |
|
кишечника |
Яйцо-реципиент
Ядро из клетки кишечника
Бластула Бластула Отсутствие дробления
Головастик
Аномальный
зародыш
5 
 Взрослая лягушка
Взрослая лягушка  (линия с одним ядрышком)
(линия с одним ядрышком)
Схема пересадки ядер из кишечника головастика в неоплодотворенное яйцо лягушки Xenopus laevis (Из: Г¸рдон, 1970, стр. 20). Ядро-реципиент, помеченное наличием двух ядрышек, разрушается в результате облучения ультрафиолетовыми лучами (1), затем у головастика выделяют кишечник (2) и берут клетки эпителиального слоя. Одиночную эпителиальную клетку засасывают в микропипетку, при этом клеточная оболочка разрушается (3), высвобождая ядро. Ядро клетки кишечника пересаживают в подготовленное яйцо (4), которое затем развивается. Около 1% яиц с пересаженными ядрами развиваются в лягушек, имеющих в ядре лишь одно ядрышко вместо обычных двух (5).
Схема опыта была аналогичной той, |
энуклеированный ооцит с целой клеткой- |
|||
которую использовали Дж. Г¸рдон и его |
донором. Экспериментально полученные |
|||
сотрудники. Маркерами в данном случае |
зиготы помещали в яйцеводы самок, где они |
|||
служили масть овец и различные |
начинали дробиться и развивались в морулы, |
|||
микросателлиты в составе ДНК. Ооциты |
которые и были пересажены в матки |
|||
выделяли из овецшотландской черномордой |
черномордых |
îâåö. |
Èç |
277 |
породы, а донорные клетки были выделены |
экспериментально полученных зигот только |
|||
из вымени овец беломордой породы Финн |
одна прошла все стадии развития вплоть до |
|||
Дорсетт. После этого с помощью |
рожденияягн¸нка,которыйбылбеломордым |
|||
электрического импульса сливали |
(Рис. 15.6). (Детали экспериментов можно |
|||
|
найти в статье: Wilmut et al., 1997). |
|
||
|
|
|
|
|
360

Генетика развития |
|
|
|
|
|
Глава 15 |
|||
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
||
Рисунок 15.6 |
|
|
|
|
Дополнение 15.1 |
|
|
||
|
|
|
|
|
История трансплантаций ядер уходит |
||||
|
|
|
|
|
в далекие 1940-е годы, когда российский |
||||
|
|
|
|
|
эмбриолог Г.В. Лопашов разработал метод |
||||
|
|
|
|
|
пересадки ядер в яйцеклетку лягушки. В |
||||
|
|
|
|
|
июне 1948 года он отправил в “Журнал |
||||
|
|
|
|
|
общей биологии” статью, написанную по |
||||
|
|
|
|
|
материалам собственных экспериментов. |
||||
|
|
|
|
|
Однако, в августе 1948 года состоялась |
||||
|
|
|
|
|
печально известная сессия ВАСХНИЛ и |
||||
Овечка (на рисунке слева), развившаяся из |
|
|
наборэтойстатьибылрассыпан.РаботуГ.В. |
||||||
|
|
Лопашова |
забыли, |
à â 1952 |
ãîäó |
||||
клетки молочной железы, взятой от овцы |
|
|
|||||||
|
|
американские эмбриологи Р.Бриггс и Т.Дж. |
|||||||
беломордой породы и трансплантированной |
|
|
|||||||
|
|
Кингвыполнилисходныеопыты.Наиболее |
|||||||
в овцу черномордой породы (справа) (Из: |
|
|
|||||||
|
|
успешные эксперименты были проведены |
|||||||
Wilmut et al., 1997). |
|
|
|
||||||
|
|
|
Дж. Гердоном в 1962 году. |
|
|||||
|
|
|
|
|
|
||||
Тот факт, что овечка выросла из |
|
||||||||
|
Целые растения вырастают и из |
||||||||
яйцеклетки с ядром из взрослого животного, |
|
||||||||
отдельных клеток, входящих в состав |
|||||||||
доказывает отсутствие необратимых |
|
каллуса (см. Першина, 2000). |
|
||||||
модификаций |
генетического материала в |
|
|
|
|
|
|||
ходе нормального развития. |
|
|
Литература к разделу 15.3. |
||||||
К 2000 году клонированные мыши и |
Гердон Дж. Пересадка ядер и клеточная |
||||||||
овцы получены после пересадки ядер из |
|
дифференцировка. В кн. “Молекулы и |
|||||||
культуры клеток в энуклеированные |
|
||||||||
|
клетки”, вып. 5, Москва, Мир, 19-37, 1970. |
||||||||
|
|
|
|
|
|||||
яйцеклеткиэтихживотных. |
|
|
Корочкин Л.И. Клонирование животных. |
||||||
Создан |
проект выращивания |
|
Соросовский образовательный журнал |
||||||
клонированныхживотных,которыемоглибы |
4: 10-16, 1999. |
|
|
||||||
быть источником органов для пересадок |
Корочкин Л.И. Введение в генетику развития. |
||||||||
больным людям. Здесь |
наиболее |
|
Москва, Наука, 24-40, 1999. |
|
|||||
привлекательным объектом являются |
Першина |
Ë.À. |
Культивирование |
||||||
свиньи. Планируется в ядре культивируемой |
изолированных |
клеток и |
тканей |
||||||
|
растений. Часть 1. Новосибирск, Изд-во |
||||||||
клетки свиньи вначале инактивировать гены, |
|
||||||||
Новосиб. госуд. универ., 1-44, 2000. |
|||||||||
ответственные за включение в клетки |
|||||||||
McCreath K.J. et.al., Production of gene-targeted |
|||||||||
полисахаридов, которые организмом |
|
sheepbynucleartransferfromculturedsomatic |
|||||||
человека опознаются в качестве чужих и |
|
||||||||
|
cells. Nature 405: 1066-1067, 2000. |
||||||||
вызывают иммунный ответ. Получены |
|
||||||||
Wakayama T, Rodrigues I., Perry A.C.F., |
|||||||||
первые результаты по получению овцы в |
|
Yanagimachi R., Mombaerts P. Mice cloned |
|||||||
результате |
трансплантации |
ÿäðà ñ |
|
|
|
96 |
|||
искусственно внесенным в него геном α 1(I) |
|
from embryonic stem cells. PNAS |
: 14984- |
||||||
14989, 1999. |
|
|
|||||||
проколлагена (см. McCreath et. al., 2000). |
Wilmut I., Schnieke A.E., McWhir J., Kind A.J., |
||||||||
В исследованиях растений проблема |
|
CampbellK.H.S.Viableoffspringderivedfrom |
|||||||
тотипотентности не возникала, т.к. всем |
|
|
fetal and adult mammalian cell. Nature 385: |
||||||
|
810-813, 1997. |
|
|
||||||
известно, что кусочки листа (т.е. |
|
|
|||||||
|
|
|
|
|
|||||
соматические клетки) бегонии дают новые |
15.4. Детерминация |
|
|||||||
растения с цветами и семенами, точно также |
|
Известно, чтокаждая клетка находится |
|||||||
корневые побеги малины ежегодно дают |
|
||||||||
в некотором детерминированном состоянии. |
|||||||||
начало новым растениям, а всем известный |
|||||||||
картофель размножается клубнями. |
Детерминация представляет собой важное |
||||||||
|
|
|
|
|
|||||
событие, в результате которого клетки с
361

Глава 15 |
|
|
|
|
|
|
|
|
Генетика развития |
|||
|
|
|
|
|
|
|||||||
одинаковым набором генов начинают |
|
|
|
|
|
|||||||
|
Рисунок 15.7 |
|
|
|
||||||||
различаться по своим внешним признакам, |
|
|
|
|
|
|||||||
или фенотипу. Много серьезных вопросов |
|
|
|
|
|
|||||||
стоит перед исследователями: на какой |
|
|
|
|
|
|||||||
стадии развития зародыша происходит эта |
|
|
|
|
|
|||||||
детерминация? Насколько устойчиво такое |
|
|
|
|
|
|||||||
состояние клетки? Наследуется ли это |
|
|
|
|
|
|||||||
состояние всеми клетками, происходящими |
|
|
|
|
|
|||||||
îò |
детерминированной |
клетки- |
|
|
|
|
|
|||||
предшественника? Можно ли изменить |
|
|
|
|
|
|||||||
детерминированность и переключить эти |
|
|
|
|
|
|||||||
клетки на развитие в новых направлениях? |
|
|
|
|
|
|||||||
|
Для ответов на эти вопросы |
|
|
|
|
|
||||||
швейцарский генетик Э. Хадорн (Рис. 15.7) |
|
|
|
|
|
|||||||
использовал плодовую мушку дрозофилу. |
|
|
|
|
|
|||||||
Известно,чтовразвитиивысшихнасекомых, |
|
|
|
|
|
|||||||
в том числе мух, происходит интересное |
|
|
|
|
|
|||||||
разделение клеток по их функциям. Клетки |
|
|
|
|
|
|||||||
одного типа начинают дифференцироваться |
|
|
|
|
|
|||||||
спервыхэтаповэмбриональногоразвития,из |
|
|
|
|
|
|||||||
них образуется тело личинки насекомого со |
|
|
Эрнст Хадорн |
|
||||||||
всеми его органами. Клетки другого типа |
|
|
1902-1976 |
|
||||||||
обособлены, |
îíè |
составляют так |
|
|
||||||||
глазной имагинальный диск, в брюшке |
||||||||||||
называемыеимагинальныедискиилизачатки |
личинки-хозяина развивается полностью |
|||||||||||
взрослых |
орагнов. |
Õîòÿ |
клетки |
сформировавшийся глаз. |
|
|||||||
имагинальных дисков находятся в контакте с |
|
В результате многих опытов Э. Хадорн |
||||||||||
соседними |
|
дифференцирующимися |
|
|||||||||
|
обнаружил, что каждый имагинальный диск |
|||||||||||
клетками, они находятся в эмбриональном |
представляет собой своего рода мозаику из |
|||||||||||
состоянии в течение всего личиночного |
различных групп клеток; например, из одних |
|||||||||||
периода.Вэтовремяониделятся.Впроцессе |
участков мужского генитального диска |
|||||||||||
метаморфоза |
|
значительная |
часть |
образуется семяизвергательный канал, из |
||||||||
личиночных органов рассасывается (или |
других - различные элементы мужского |
|||||||||||
лизируется). Одновременно с этим клетки |
полового органа, из третьих - анальные |
|||||||||||
имагинальных дисков утрачивают свое |
пластинки и задняя кишка. Следовательно, |
|||||||||||
эмбриональное |
состояние, |
îíè |
будущее |
разнообразие |
клеток |
|||||||
дифференцируются, |
превращаясь в |
детерминированоуженаличиночнойстадии. |
||||||||||
специализированные ткани имаго (взрослой |
Неожиданные |
результаты |
áûëè |
|||||||||
мухи). Из каждого диска образуется |
||||||||||||
получены |
после |
трансплантации |
||||||||||
отдельная часть тела насекомого. Например, |
имагинальныхдисковсразувовзрослыхмух. |
|||||||||||
для каждой из шести будущих ног |
Клетки неограниченно делились и |
|||||||||||
существует |
отдельный диск, |
голова |
разрастались. Если бы развитие этих |
|||||||||
образуется из трех пар дисков. |
|
имагинальных дисков происходило в |
||||||||||
|
Имагинальные дискиможно извлечь из |
|||||||||||
|
нормальной личинке, то эти клетки |
|||||||||||
тела личинки и пересадить в полость тела |
прекратили бы деление с началом |
|||||||||||
другой личинки. Когда личинка-хозяин |
метаморфоза: под влиянием гормона |
|||||||||||
превращается в куколку, трансплантант |
насекомых экдистерона они бы начали |
|||||||||||
дифференцируется в соответствующий |
дифференцироваться в такие структуры |
|||||||||||
орган. Например, если трансплантирован |
взрослых особей, как щетинки, волоски, |
|||||||||||
362

Генетика развития |
Глава 15 |
|
|
коготки и т.д. Неограниченный рост этих клеток, пересаженных в брюшко взрослых мух, продолжался более 6 лет. Поскольку муха дрозофила живет около месяца, размножающиеся клетки пересаживали в новуюмухучерезкаждые2недели.Приэтом клетки перенесли свыше 160 пересадок. Хотя трансплантанты жили во взрослых мухах годами, они сохранили свой исходный эмбриональный характер и не дифференцировались. Если же извлечь немного трансплантанта и ввести в личинку, эти клетки претерпевали метаморфоз и нормальнодифференцировалисьвструктуры взрослого организма. При этом, если нескольколетназаддлятрансплантациибыли взяты крыловые имагинальные диски, при обратнойтрансплантациитакжеформировалось крыло.Такимобразом,состояниедетерминации можетвоспроизводитьсядлительноевремябез каких-либоизменений.Оченьпродолжительное времяэтосвойстводетерминированныхклеток передаетсяблагодарясвоегорода“клеточной наследственности”, или определенного состояния ядерно-цитоплазматических отношений (наличия эпигенетических факторов).
В некоторых случаях нормальное состояние детерминированности в опытах Э. Хадорна резко изменялось. Очень редко, из клетокгенитальногоимагинальногодискапосле длительного размножения в брюшке имаго, формировалисьорганыголовыиликонечности, т.е. клетки больше не дифференцировались в соответствие с детерминацией их предков - произошла трансдетерминация. Вновь приобретенное трансдетерминированное состояние в дальнейшем также передается за счет клеточной наследственности очень продолжительноевремя.
Явлениетрансдетерминации,такжекаки результатытрансплантацииядер,полученные Дж. Гердоном, свидетельствуют, что в основе дифференцировки не лежит необратимое изменения генов,темболееихпотеря.
Очевиднотакжеито,чтовосновелюбого детерминированного состояния лежит сбалансированная система ядерно-
цитоплазматических отношений. Как может сформироваться такая система лучше всего показывают результаты самого раннего развития.
Литература к разделу 15.4.
Хадорн Э. Трансдетерминация клеток. В кн. “Молекулы и клетки”, вып. 5, Москва, Мир, 54-61, 1970.
Hadorn E. Transdetermination in cells. Scientific American November 1968: 110-120.
15.5. Раннее эмбриональное развитие дрозофилы
Еще в 1950-е годы сформировалось представлениеоморфогенахкаковеществах, индуцирующих образование определенных частейтела.Предполагали,что“этивещества диффундируютчерезтканьиихраспределение диктует тот или иной путь развития клетки”. Позднее теория морфогенов получила значительное развитие. По современным представлениям, морфоген выделяется из локального источника и во время последующей диффузии в ткани образуется градиентегоконцентрации.Вкаждойгруппе клетоксуществует свойнабориконцентрация морфогенов, т.е. своя информация о последующемразвитии-этото,чтогенетики развитияназываютпозиционнойинформацией.
Лучше всего изучены градиенты морфогенов,образующиесявразвивающемся яйцедрозофилы.
Известно,чтоудрозофиляйцосозревает
âособой камере - фолликуле. Эта камера содержит ооцит - созревающее яйцо и 15 огромныхпитающихклеток,функциякоторых - синтезироватьпродукциюиперекачиватьее
âооцит. В них функционируют т.н. “гены с материнским эффектом”, т.е. такие гены, которыефункционируютвпитающихклетках ооцитов - в организме матери еще до оплодотворения яйца сперматозоидом и информация от нихпередаетсявооцит.Один из таких генов - bicoid (bcd). Самки, гомозиготные по мутации bcd, откладывают яйца, в которых нормальные эмбрионы не развиваются, даже если эти яйца
363

Глава 15 |
|
|
|
|
|
|
Генетика развития |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
оплодотворены спермием, содержащим |
|
|
|
|
|
|
|
|
|
|
|
|
|
Рисунок 15.8 |
|
|
|
|
|
|
|
|
|||
нормальный аллель гена bcd. Совершенно |
|
|
|
|
|
|
|
|
|
|
|
|
очевидно, что весь продукт этого гена, |
|
à |
|
|
|
|
|
|
|
|
|
|
необходимый для развития, синтезируется у |
|
|
|
|
|
|
|
|
|
|
|
|
материиоткладываетсявяйце. |
|
|
|
|
|
|
|
|
|
|
|
|
Оказывается, что белки, кодируемые |
|
|
|
|
|
|
|
|
|
|
|
|
генами,функционирующимивходесозревания |
|
á |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|||
яйца и транспортируемые туда из питающих |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
клеток, распределяются по оси яйца, образуя |
|
|
|
|
|
|
|
|
|
|
|
|
градиенты,характерныедляпродуктовкаждого |
|
â |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
гена. На Рис. 15.8 показано распределение |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
||
продуктагенаbcdвпределахяйца.Онзанимает |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
строгоопределенныйучасток.Чтобыпродукт |
|
|
|
|
|
|
|
|
|
|
|
|
bcd занялэтоместо,нужно,чтобыпоработали |
|
ã |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
идругиегены,вслучаемутацийкоторыхданный |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
||
продукт распределяется неправильно. Так, в |
|
|
|
|
|
|
|
|
|
|
|
|
|
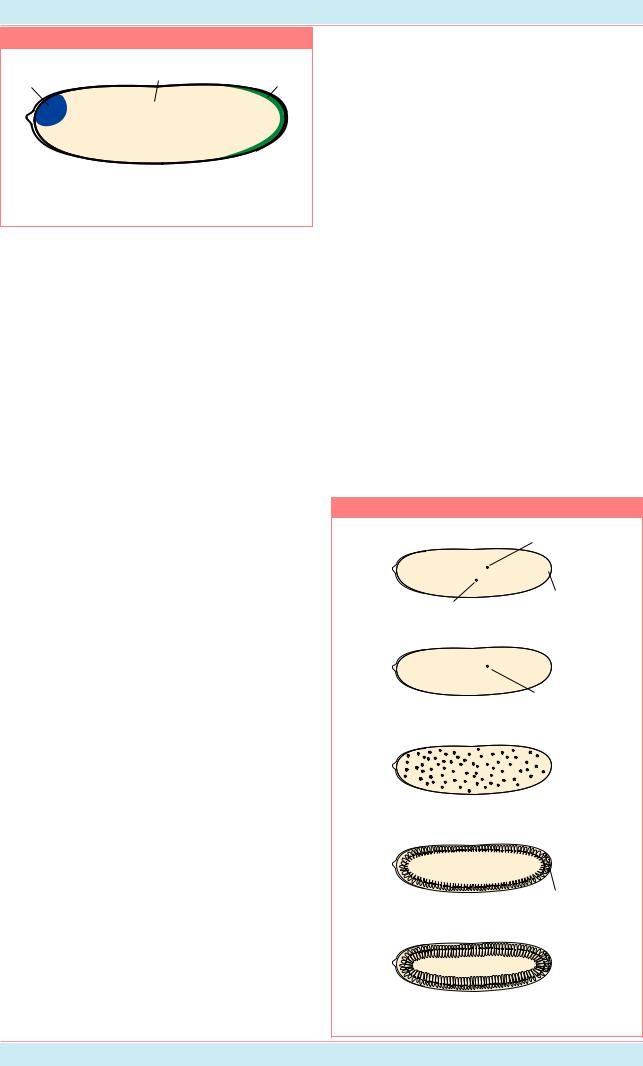
Распределение по длине яйца дрозофилы |
|||||||||||
нормальномяйцеРНКгенаbcdрасполагаетсяв |
|
|||||||||||
|
матричной РНК, считанной с гена bicoid в |
|||||||||||
узколокальномучастке(Рис.15.8a).Однако, у |
|
|||||||||||
|
нормальной линии (синее пятно) (а), у |
|||||||||||
рядамутантовраспределениеэтойРНКвяйце |
|
|||||||||||
|
мутантов exuperantia (б), swallow (в) и staufen |
|||||||||||
сильно изменено: в результате мутации гена |
|
|||||||||||
|
(г) (показаны штриховкой) (Из: Lawrence, 1992, |
|||||||||||
exuperantia РНК гена bcd более или менее |
|
|||||||||||
|
p. 31). |
|
|
|
|
|
|
|
|
|||
равномерно распределена по всему яйцу с |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
||
каждая из этих РНК ещ¸ и распределяется по |
||||||||||||
небольшимградиентомотпереднегополюсак |
||||||||||||
своим местам в яйце в результате активности |
||||||||||||
заднему. У мутантов swallow градиент этого |
||||||||||||
морфогенавыраженсильнее,т.е.распределение |
других генов, совершенно очевидно сколь |
|||||||||||
огромно число генов, участвующих в |
||||||||||||
егоближекнормальному:впереднейчастияйца |
||||||||||||
формированиияйца. |
|
|
|
|
||||||||
выявлено большое скопление РНК bcd и |
|
|
|
|
||||||||
|
Присозреваниияйцаворганизмематери |
|||||||||||
некоторое е¸ количество распределено в |
|
|||||||||||
происходит |
формирование |
четырех |
||||||||||
остальнойцитоплазме(Рис.15.8б,в). |
независимых систем: 1. Передне-заднего |
|||||||||||
Совсем близкое к норме, но все ещ¸ |
||||||||||||
градиента белков(РНК)гена bcd,2.Градиента |
||||||||||||
ненормальное, распределение продукта bcd |
||||||||||||
распределения |
белка |
ãåíà |
nanos, |
|||||||||
обнаружено у мутантов staufen (Рис. 15.8г): у |
||||||||||||
расположенного в задней части яйца и |
||||||||||||
них РНК bcd вообще не переходит в заднюю |
||||||||||||
необходимого для развития брюшка мухи. 3. |
||||||||||||
частьэмбриона. |
||||||||||||
Формирование терминирующей системы - |
||||||||||||
Таким образом, РНК считывается с гена |
||||||||||||
bcd ещ¸ в питающих клетках ооцитов в |
градиентабелкагенаtorso, расположенногона |
|||||||||||
обоих полюсах яйца и необходимого для |
||||||||||||
материнском организме и поступает в |
||||||||||||
яйцеклетки. Затем с помощью продуктов |
определенияголовнойихвостовойчаститела. |
|||||||||||
4.Дорзо-вентральнаясистема,котораязависит |
||||||||||||
других генов, в данном случае это гены |
||||||||||||
от активирования рецепторного белка, |
||||||||||||
exuperantia, swallow è staufen, ÐÍÊ bcd |
||||||||||||
кодируемогогеномToll (часть из них показана |
||||||||||||
занимаетопредел¸нноеположениевцитоплазме |
||||||||||||
íà Ðèñ. 15.9). |
|
|
|
|
|
|
|
|
|
|||
яйца, т.е. созда¸тся определ¸нный градиент в |
|
|
|
|
|
|
|
|
|
|||
|
 ñâîþ |
|
|
очередь, |
после |
занятия |
||||||
распределении данного морфогена. В случае |
|
|
|
|||||||||
правильногоположениявяйце,продуктытаких |
||||||||||||
мутациилюбогоизтр¸хперечисленныхгенов |
||||||||||||
генов, как bcd, вступают во взаимодействие с |
||||||||||||
распределение РНК bcd изменяется, что |
||||||||||||
другимигенами,которыеактивируютсяпосле |
||||||||||||
приводитксерь¸знейшимнарушениямразвития. |
||||||||||||
оплодотворения и образования зиготы |
||||||||||||
Известно, что в яйцо поступает РНК, |
||||||||||||
(зиготические |
гены). Белки гена bcd |
|||||||||||
считаннаясогромногочислагенов.Поскольку |
||||||||||||
связываются с контролирующими районами |
||||||||||||
|
||||||||||||
364
Генетика развития Глава 15
Рисунок 15.9 |
|
|
|
|
|
блокированию транскрипции (если белок |
|||
|
|
hunchback |
|
|
|
является репрессором). |
|
||
bicoid |
|
torso |
|
nanos |
НаРис.15.11изображенфрагмент(около |
||||
|
|
|
|
||||||
|
|
|
|
|
|
|
700 п.н.) регуляторной части гена eve (even- |
||
|
|
|
|
|
|
|
skipped), который контролирует развитие |
||
|
|
|
|
|
|
|
правильной сегментации тела дрозофилы. |
||
Распределение |
морфогенов |
ÐÍÊ |
ïî |
Видно, что мотивы |
нуклеотидов, |
||||
связывающихактивирующиебелкигеновbcd |
|||||||||
продольной оси яйца дрозофилы (Из: Lawrence, |
|||||||||
и hb, часто перекрываются с мотивами, на |
|||||||||
1992, p. 42). |
|
|
|
|
|
||||
|
|
|
|
|
которые садятся белки, подавляющие |
||||
|
|
|
|
|
|
|
|||
зиготическихгеновиактивируютих.Понятно, |
транскрипцию(геныKr иgt). |
||||||||
чтоклетка,возникающаявобластилокализации |
Эти данные свидетельствуют о том, что |
||||||||
морфогенаbcd,будетиспытыватьеговлияние, |
расположение белков, синтезированных в |
||||||||
иразвитиепойдетвопредел¸нномнаправлении. |
материнскоморганизме,вопределеннойчасти |
||||||||
Если же клетка расположена в задней части |
яйца (см. Рис. 15.9) имеет первостепенное |
||||||||
эмбриона, где морфогена bicoid нет (см. Рис. |
значениедляпроцессаактивированиягеновв |
||||||||
15.8), она будет развиваться в другом |
уже начавшем развитие эмбрионе. Ясно, что |
||||||||
направлении. Таким образом, набор |
ген eve будет функционировать в той части |
||||||||
определенных |
белков, |
накопленных |
эмбриона, в которой содержится много |
||||||
цитоплазмой к данной стадии развития, |
материнских белков bcd и hb, и не будет |
||||||||
способен активировать определенный набор |
функционировать в клетках, содержащих |
||||||||
генов, благодаря чему либо поддерживается |
избыток белков Kr и gt. |
|
|||||||
данноедифференцированноесостояние,либо |
Рисунок 15.10 |
|
|||||||
развитиепродвигаетсядальше. |
После того |
|
|||||||
Оплодотворенное яйцо |
|
||||||||
|
|
|
|
|
|
|
|
||
как градиенты в яйце созданы, происходит |
с двумя родительскими ядрами |
Пронуклеус самца (N) |
|||||||
оплодотворение и начинается дробление |
Передний |
Задний |
|||||||
|
|
|
|
|
|
|
|||
зародыша (Рис. 15.10), в результате чего |
конец |
конец |
|||||||
ÿéöà |
ÿéöà |
||||||||
образуетсяоднослойнаябластодерма.Каждая |
|
Полярная |
|||||||
Пронуклеус самки (N) |
цитоплазма |
||||||||
клеткавнейзанимаетопределенное положение |
Слияние родительских ядер |
|
|||||||
по отношению |
ê |
сформировавшимся |
и образование зиготы |
|
|||||
|
|
||||||||
градиентам, т.е. обладает определенной |
|
|
|||||||
позиционной информацией. Морфогены |
|
|
|||||||
взаимодействуютсрегуляторнымиучастками |
|
Ядро зиготы |
|||||||
Образование синцития в результате |
2N |
||||||||
генов, |
активирующихся |
ó |
зигот |
(ò.å. |
|
||||
9 последовательных делений ядер |
|
||||||||
|
|
|
|
|
|
|
|
||
зиготическихгенов). |
|
|
|
|
в общей цитоплазме |
|
|||
|
|
|
|
|
|
||||
Какимобразомбелковыйпродуктодного |
|
|
|||||||
гена может взаимодействовать с другим |
|
|
|||||||
геном?Какизвестнорегуляторныечастигенов |
Ядра мигрируют к стенкам яйца |
|
|||||||
содержатспецифическиегруппынуклеотидов |
и делятся еще 4 раза, образуя |
|
|||||||
однослойный мешок |
|
||||||||
(мотивы),имеющиесродствокопределенным |
|
|
|||||||
сочетаниям аминокислот (доменам) в |
|
|
|||||||
молекулах белков. Посадка различных |
|
Полярные клетки |
|||||||
белковых факторов на соответствующие |
Бластодерма из 4000 клеток. |
(зачатки клеток |
|||||||
Завершение образования |
зародышевого пути) |
||||||||
мотивы в регуляторных областях генов |
|||||||||
клеточных мембран |
|
||||||||
|
|
|
|
|
|
|
|
||
приводиткизменениямконфигурацииДНП(см. |
|
|
|||||||
более подробно раздел 7.6) и началу |
|
|
|||||||
транскрипции кодирующей части гена (если |
Эмбриональное развитие дрозофилы (Из: |
||||||||
белок |
является |
активатором) |
èëè |
||||||
Russell, 1998, p. 570). |
|
||||||||
|
|
|
|
|
|
|
|
||
365
Глава 15 |
Генетика развития |
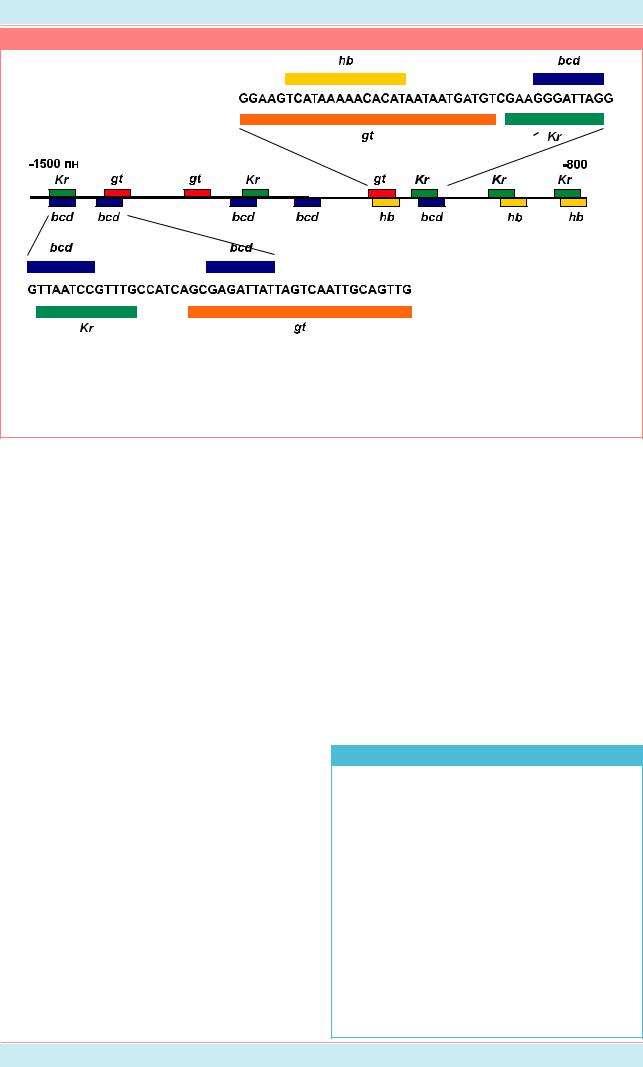
Рисунок 15.11 |
|
Часть регуляторного участка гена eve длиной 700 пар нуклеотидов (от -800 до -1500 пн) (Из: |
|
Lawrence, 1992, p. 73). На рис. а нарисована в масштабе нить ДНК, выше и ниже которой |
|
соответствующими символами изображены участки посадки белков, кодируемых генами Kr, |
|
gt, bcd и hb. На рисунках б и в показаны последовательности нуклеотидов, связывающие |
|
данные белки. |
|
После образования бластодермы и включения зиготических генов начинается формирование сегментального плана строения тела (Рис. 15.12). Дело в том, что тело взрослой особи насекомых (имаго) состоит из ясно выраженных сегментов. В бластодерме сегментация выявляется лишь по выявлению продуктов определенных генов,сегментычетко не определеныи носят название парасегментов. На более поздних стадиях развития сегменты хорошо обособляются друг от друга и легко обнаруживаются (Рис. 15.12). Эмбриональные сегменты дают начало сегментам взрослой мухи.
Сегментный тип организации свойственен не только двукрылым насекомым, но и многим животным. Даже у человека на некоторых этапах развития выявляются сомиты - первичные сегменты тела, на которые разделяется в ходе зародышевого развития мезодерма.
Сегменты формируются в результате действия генов сегментации, 25 из которых охарактеризовано к настоящему времени. В результате мутаций генов сегментации изменяется число и расположение сегментов. Гены сегментации разделяются
на три большие группы, выделяемые по их мутантному проявлению в фенотипе (Рис. 15.13): Gap-гены, Pair-rule гены и гены сегментной полярности (Таблица 15.1). Мутации группы Gap (от английского слова gap – брешь) приводят к потере нескольких прилежащих сегментов тела, в результате чего в рисунке сегментации образуется пустота или брешь (Рис. 15.13). Мутации в группе генов pair rule (или генов “правила парности”) приводят к утрате одного и того же фрагмента в каждом втором сегменте. У мутантов по генам сегментной полярности определенные части сегментов заменены
Таблица 15.1
Список генов, участвующих в контроле сегментации тела дрозофилы (Из: Корочкин, 1999, стр. 94).
Gap-ãåíû |
Pair-rule ãåíû |
Гены сегментной |
|
полярности |
|||
|
|
||
|
|
|
|
Kruppel |
hairy |
engrailed |
|
|
|
|
|
knirps |
even-skipped |
wingless |
|
|
|
|
|
hunchback |
runt |
cubitus interruptus |
|
|
|
|
|
giant |
fushi-tarazu |
hedgehog |
|
|
|
|
|
tailless |
odd-paired |
fused |
|
|
|
|
|
huckebein |
odd-skipped |
armadillo |
|
|
|
|
|
|
sloppy-paired |
patched |
|
|
|
|
|
|
paired |
gooseberry |
|
|
|
|
366