

Подготовка у универсиаде 2012 / Генетика (Жимулев) / 18ver7
.pdfГены в популяциях |
Глава 18 |
|
|
Глава 18. ГЕНЫ В ПОПУЛЯЦИЯХ
18.1. Популяции и генофонд
Любойвидживыхсуществраспространен по занимаемому им ареалу не сплошь, а в той или иной мере обособленными друг от друга совокупностямиособей-популяциями.Условия жизнивразныхместахареаланеоднородны,и особи данного вида концентрируются преимущественновнаиболееблагоприятных для них участках ареала. Однако, не всякая изолированная группа особей представляет популяцию,т.к.впределахгруппыособичасто тоже распространены неравномерно, образуя относительнонемногочисленныегруппировки, сохраняющиеся лишь в течение короткого времени(одного-двухпоколений).
Длядальнейшегопониманияважноиметь представление о панмиктической и подразделеннойпопуляциях.Впанмиктической популяции слагающие ее особи свободно скрещиваютсядругсдругом.Подразделенная популяция представлена совокупностью субпопуляций,испытывающиходновременное воздействиеслучайногодрейфа,миграциигенов иестественногоотбора.
Наиболее четкое определение реально существующейпопуляциидалиН.В.ТимофеевРесовский, А.В. Яблоков и Н.В. Глотов в 1973 году для видов, размножающихся половым путем при перекрестном оплодотворении: популяция-этосовокупностьособейданного вида, в течение длительного времени (большого числа поколений) населяющая определенный ареал и состоящая из особей, имеющих возможность скрещиваться друг с другом и отделенная от таких же соседних совокупностей одной из форм изоляции (пространственной,сезонной,физиологической илигенетической).
Реальные популяции ограничены по численности, кроме того, генетически эффективнаячисленность(Ne)популяциивсегда исущественноменьшеееобщейчисленности (Nt)ужепотойпростойпричине,чтостаршиеи младшие возрастные группы исключены из процесса воспроизводства. У человека,
например, если обратиться к сельскому населению,величинаNeсоставляетпримерно треть от величины Nt. У природных видов отношение Ne/Nt может быть еще меньше, порядка0.1-0.2.
При изучении процесса эволюции популяцийважноезначениеимеетпредставление огенофонде.Генофондом(genepool)называют совокупность генотипов всех особей популяции. Для диплоидных организмов генофондпопуляции,насчитывающейNособей, состоит из 2N гаплоидных геномов. Таким образом, генофонд популяции из N особей включаетвсебяпо2Nгеновкаждоголокусаи N пар гомологичных хромосом. Исключение составляютполовыехромосомыисцепленные с полом гены, представленные в каждом гетерогаметноморганизмеводномэкземпляре.
18.2. Закон Харди-Вайнберга
При переходе от индивидуальных скрещиваний, связанных с анализом наследованиявсемьях,кгенетикепопуляций, где единой теорией необходимо охватить последствия от множества свободных скрещиваний, встает ряд новых проблем, в частности, каким образом в популяциях сохраняются как доминантные, так и рецессивные гены? Почему доминантные аллели не вытесняют рецессивные? На этот вопрос в 1908 году попытались ответить английскийматематикДж.Х.Харди(G.H.Hardy) инемецкийврачГ.Вайнберг(G.Weinberg).
Работая независимо, Харди и Вайнберг показали, что генетические расщепления, которые происходят в каждом поколении у диплоидных организмов, сами по себе не изменяют общего состава генофонда (gene pool). Чтобы продемонстрировать это, они исследовали поведение аллелей в идеализированной популяции, в которой соблюдаютсяследующиепятьусловий:
1.Новые мутации в данной популяции не появляются.
2.Популяцияполностьюизолирована,т.е.нет миграцииособей-носителейгеноввпопуляцию (иммиграция)иизпопуляции(эмиграция).
401

Глава 18 |
Гены в популяциях |
|
|
3.Популяциябесконечновелика,кнейможно применять законы вероятности, т. е. когда в высшей степени маловероятно, что одно случайное событие может изменить частоты аллелей.
4.Скрещиванияслучайны,т.е.происходитчисто случайное образование родительских пар - панмиксия.
p+q=1.Предположим,чтовкакой-топопуляции частотааллеляA -составляет80%,т.е.частота A-0.8,илиp=0.8.Зная,чтоаллелейтолькодва, частотааллеляaбудет0.2(q=1-p).Представим, чтоотносительныечастотыAиaодинаковыкак усамцов,такисамок,приэтомсамцыисамки скрещиваютсясовершеннослучайно.Составив решеткуПэннетта,можноподсчитатьчастоты
5.Все аллели равно влияют на результирующих генотипов (Рис. 18.1):
жизнеспособностьгамет,другимисловаминет различийврепродуктивномуспехе.Потомкиот всех возможных скрещиваний имеют равновероятнуювыживаемость.
Рассмотрев одиночный ген, имеющий только два аллеля, A и a, Харди и Вайнберг с помощьюматематическогоуравненияпоказали, чтоотносительныепропорцииаллелей Aи aв такойидеальнойпопуляциинеизменяютсяот поколения к поколению. Более того, частоты трех возможных комбинаций этих аллелей - генотипы AA, Aa и aa - не изменяются от поколения к поколению. Частоты генов находятся в состоянии равновесия по этим аллелям. Это равновесие выражается следующимуравнением:
p +2pq+q =1
В этом уравнении буква p обозначает частоту одногоаллеля,абукваq-другогоаллеля.p+q всегда должны составлять единицу, p - обозначает частоту особей, гомозиготных по одному аллелю, q - частота особей, гомозиготных по другому аллелю и 2pq - частота гетерозигот. Как видно, соотношение гомо - и гетерозиготных генотипов в ней соответствует коэффициентам разложения биномаНьютона.
Чтобы понять, как Харди и Вайнберг вывелисвоеуравнениеипродемонстрировали равновесие частот аллелей и генотипов в популяциях при условии выполнения пяти вышеупомянутых условий, можно более подробно рассмотреть ситуацию с единственным геном, имеющим только два аллеля, A и a. Важно проследить, как относительные пропорции аллелей A и a передаютсяотодногопоколениякдругому.Как былоуказановыше,еслиестьтолькооднапара аллелей, p è q вместе должны дать единицу:
64% - AA, 32% - Aa è 4% - aa.
Можноподсчитатьчастотыипоформуле Харди-Вайнберга: p + 2pq + q = 1
p =0.8× 0.8=0.64 (частота генотипов AA), 2pq=2× 0.8× 0.2=0.32(частотагенотиповAa), q =0.2× 0.2=0.04 (частота генотипов aa).
Чтожеслучилосьсчастотамидвухаллелей в генофонде в результате этого скрещивания? ЧастотаAAсоставляет0.64,плюсполовинаот 0.32 (генотипы Aa), т.е.получается 0.64+0.16=0.8 ичастотааллеляAнеизменилась. Считаяаналогичнымобразом,получаемчастоту аллеля a=0.04+ 0.16 (половина от 0.32) = 0.2. Отсюда вытекает, что частота аллеля a также не изменилась. Таким образом, в идеализированной популяции, в которой выполняютсяуказанные5условий,ничастота аллелей,ничастотагенотиповнеизменяетсяот поколениякпоколению.
Различные диплоидные комбинации множественныхаллелеймогутдаватьразличные переходные фенотипы. Уравнение ХардиВайнбергаприменимоикэтимситуациям,хотя оно становится более сложным. Например, частоты генотипов для трех аллелей
Рисунок 18.1 |
|
|
|
|
|
|
|
)=0.8 |
|
==0.2 |
|
|
|
|
|
||
|
|
|
|
|
|
)=0.8 |
)) |
|
)= |
|
|
0.64 |
|
0.16 |
|
||
|
|
|
|
||
|
|
|
|
|
|
==0.2 |
)= |
|
== |
|
|
0.16 |
|
0.04 |
|
||
|
|
|
|
||
|
|
|
|
|
|
Результаты случайного скрещивания в популяции, в которой частота (F) аллеля A равна 0.8 и частота аллеля a (G) равна 0.2. Принимается, что ген не сцеплен с полом и что аллели встречаются у самок и самцов с одинаковой частотой.
402
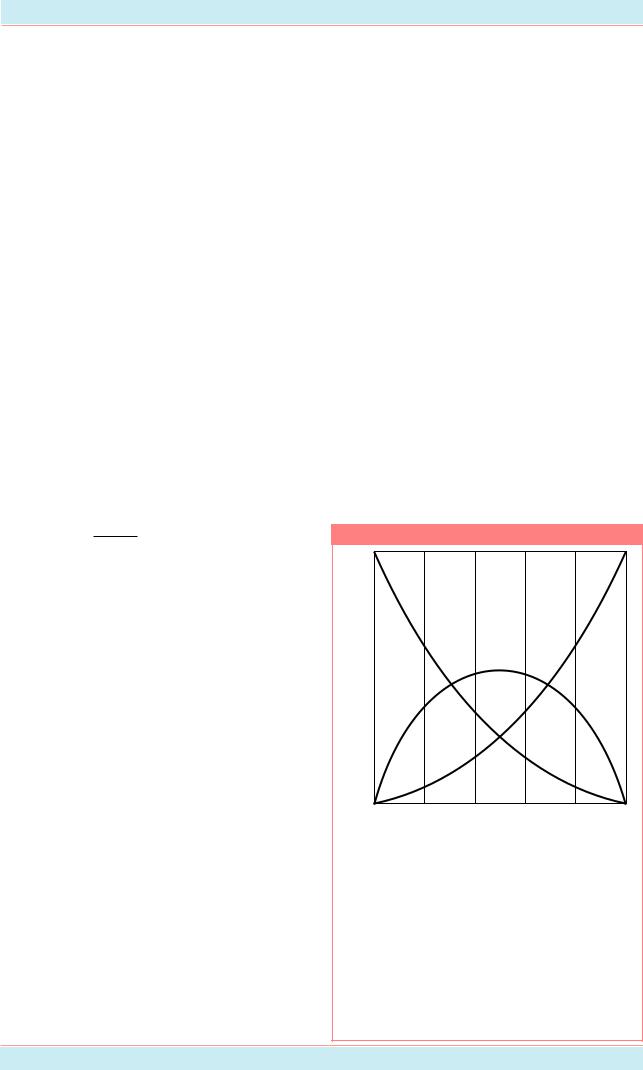
Гены в популяциях
описываются следующим выражением: (p+q+r) =1, ãäå r - частота третьего аллеля.
Уравнение позволяет оценивать изменения, происходящие в популяциях, определять их величину и направление. Если удастсянайтивпопуляциигомозиготныхособей, можноподсчитатьчастотуэтогоаллеля,азатем ичастотыостальныхгенотипов.Еслипровести эту работу в нескольких поколениях, можно увидеть, какие процессы идут в генофондах популяций,изатемискатьпричину.
Какужеуказываливыше,правилоХардиВайнберга выполняется только в том случае, если выполняются все пять условий, характеризующихидеальнуюпопуляцию.Если не выполняется хотя бы одно из них, частоты аллелей начнут изменяться. Представим, что пару аллелей A и a, один из которых - a - у гомозигот вызываетнекоторыеповреждения, несколько уменьшающие вероятность выживанияилиоставленияпотомства.Можно подсчитатьчислоособей-носителейгенотипов aa в этой популяции, допустим 1 на 10000, другими словами q=1/10000=0.0001. Таким образом, q= √ 0.0001 èëè 0.01. Åñëè q=0.01, òî p=0.99, à 2pq=0.0198 или почти 0.02. Таким образом, 2% популяции - одна особь на 50 - можетбытьгетерозиготнымносителемаллеля a.Еслисделатьэтотжеанализчерез5летинайти частоту q=0.009, а еще через 5 лет - 0.008. Другимисловами,медленноосуществляются эволюционные изменения. Частота одного аллеля уменьшается, частота другого увеличивается(Рис.18.2).
18.3. Факторы генетической эволюции в популяциях
Довольно скоро стало ясно, что правило Харди-Вайнберга неприменимо к реально существующим популяциям, на которые оказываютдавлениемногочисленныефакторы.
Â1926 году была опубликована работа
Ñ.Ñ.Четверикова, в которой впервые были показаны закономерности возникновения мутаций,ихраспространениевнутрипопуляций, каквусловияхсвободногоскрещивания,таки приискусственнойизоляции,атакжевследствие плейотропии.
Глава 18
Прямая взаимосвязь между степенью генетической изменчивости популяции и скоростью эволюции под действием естественного отбора была доказана математическимпутемР.А.Фишеромвработе, опубликованнойв1930году,гдеонввелпонятие приспособленности и сформулировал фундаментальную теорему естественного отбора.
Другимиисследователями,заложившими в1931-1932гг.основыновойэволюционнойи популяционной биологии, базирующейся на теориях Менделя и Дарвина, были С. Райт (S. Wright), Дж. Холдэйн (J.B.S. Haldane), Н .П. ДубининиД.Д.Ромашов(Дополнение18.1).
Многиефакторымогутизменятьчастоты аллелей в популяциях: естественный отбор, мутации,генныемиграции,генетическийдрейф инеслучайныескрещивания.
Действие естественного отбора
может приводить к тому, что носители одних генотипов оставят потомков больше, чем других,и этовызоветизменениегенетической
Рисунок 18.2
1.0
|
AA |
aa |
0.8 |
|
|
|
|
|
генотипов |
|
|
0.6 |
|
|
Частоты |
|
Aa |
|
|
|
0.4 |
|
|
|
|
|
0.2 |
|
|
|
|
0.2 |
0.4 |
0.6 |
0.8 |
1.0 |
Частота a
Взаимосвязи между частотой аллеля a в популяции и частоты генотипов AA, Aa и aa. Чем чаще встречаются генотипы AA, тем ниже частота a. Поскольку частоты AA, Aa и aa взаимосвязаны, любое изменение частоты любого аллеля приводит к соответствующим и симметричным изменениям в частотах другого аллеля и всех генотипов. По оси абсцисс - частота аллеля “а”, по оси ординат - частоты генотипов.
403

Глава 18 |
Гены в популяциях |
|
|
структуры популяции. Отбор означает дифференциальную вероятность оставления потомства разными особями или группами особей. Вероятность оставить потомство детерминируется многими свойствами организма - его жизнеспособностью, быстротой достижения репродуктивного возраста, продолжительностью репродуктивного периода, способностью к скрещиванию,плодовитостью.Совокупность этихсвойствназываетсяприспособленностью особи к условиям среды, в которой они обитают. Действие отбора на генетическое строение популяции состоит в том, что некоторые группы особей, генотипически отличающиеся от других, част ично или полностью устраняются из размножения, так что на генетическое строение будущего поколения оказывает влияние только оставшаясячастьпопуляции.Есливероятность оставленияпотомстваособямисгенотипомaa на 10% меньше, чем особями с генотипом AA или Aa, то приспособленность (W) этих двух групп записывают так: для особей AA или Aa W=1, для особей aa W=0.9. Мерилом интенсивности отбора служит разность приспособленности сравниваемых групп, называемая коэффициентом отбора (S). Для только что приведенного примера S=WAA- Waa=1.0-0.9=0.1
Мутации. Сохранение даже относительного постоянства частоты генов возможно только при допущении, что они не мутируют.Однакоизвестно,чтоэтонетак.
Мутации постоянно возникают в популяциях и тоже сдвигают равновесие аллелей.Изменчивостьвпопуляцияхвыявляют с помощью огромного количества приемов и методов,начинаяотанализацветанадкрылий божьих коровок или числа и формы полос на раковинах улиток, огромного объема данных по полиморфизму в электрофоретической подвижности ферментов, выявляемой в крахмальномгеле,доварьированияпервичной структурыДНК,какядерной,такинеядерной (напримермитохондриальной),варьирования рестрикционныхфрагментов.
Ужевпервыхисследованияхприродных популяций, проведенных под руководством С.С. Четверикова, было показано, что под внешненормальнымфенотипомдикихособей, живущихвприроде,скрываетсяудивительная картина громадной наследственной изменчивости. У нормальных по внешнему виду особей множество хромосом были носителями мутаций: летальных, полулетальных, субвитальных, мутаций, влияющих на фертильность, морфологию, окраску, длительность жизни и др. Каждая особьизпопуляцииимелапонесколькотаких мутаций.Насыщенностьпопуляциимутациями, снижающими ее приспособленность, называется генетическим грузом. Он определяется как среднее число летальных мутацийнаоднуособьизданнойпопуляции.
ЕслиаллельAмутируетсопределенной скоростью в a, то очевидно, что в генофонде популяции постепенно в ряду поколений частотааллеля A будетуменьшаться на число измененныхгенов,ачастотааллеляaвозрастет наэтужевеличину.Это,естественноприведет к нарушению равновесия в популяции, к изменениюеегенетическогосостава.
Генными миграциями называют перемещениеаллелейвпопуляциюиизнее.В результате миграций в популяцию вносятся совершенно новые аллели или значительно изменяютсячастотыужесуществующих.
Генетическийдрейф.Какужеизвестно, правилоХарди-Вайнбергавыполняетсятолько в том случае, если популяция достаточно велика. Это совершенно необходимо поскольку уравнение зависит от законов вероятностей.
Явление генетического дрейфа было описано в 1931 году одновременно и независимо друг от друга Н.П. Дубининым и Д.Д.РомашовымвСССРиС.РайтомвАнглии. Суть его состоит в том, что численнос ть популяции может оказывать существенное влияниенаеегенотипическуюструктуру.При резкомуменьшениичисленностипопуляции, напримерпримассовойгибелиособейвовремя необычно холодной зимы, в силу случайных причин могут сохраниться носители редких
404

Гены в популяциях |
Глава 18 |
|
|
уклонений.Онистанутисходнойформойпри последующем возрастании численности популяции, что приведет к их широкому распространению, по существу не обусловленному реальной селективной ценностью.
ДубининиРомашовназвалиэтоявление генетико-автоматическими процессами, а Райт - дрейфом генов.
Например,частотааллеляaвпопуляции составляет 1% . В популяции из 1 миллиона особей присутствует 20000 аллелей a (1 миллион особей имеют 2 млн. аллелей этого гена, 1% составляет 20000). Однако в маленькойпопуляции,состоящейиз50особей присутствуеттолькооднакопияаллеляa.Если особь, несущая этот аллель, не сможет оставить потомство или погибнет, не оставив потомства,аллель aбудетполностьюпотерян. Аналогичнымобразом,еслипогибнут10из49 особей, не имеющих аллеля a, его частота повысится с 1% до 1.25%. Это явление измененийвсоставегенофондаподдействием случайных обстоятельств, носит название генетического дрейфа. Можно выделить две разновидности дрейфа: эффект основателя и “бутылочное горлышко”. а) Эффект основателя. Малая популяция, которая возникает из большей, может представлять, а можетинепредставлятьгенетическийсостав этой большойпопуляции.Некоторыеаллели, редковстречающиесявбольшойпопуляции,в малойпопуляциимогутотсутствоватьсовсем или быть сверхпредставлены. В результате этого даже тогда, когда эта малая популяция начнет численно увеличиваться, она будет иметь другой генетический состав - другой генофонд по сравнению с родительской группой (популяцией). Это явление - один из типов генетического дрейфа, называется эффектом основателя.
б) “Бутылочное горлышко”. Это другой тип генетического дрейфа, он встречается тогда, когда популяция резко уменьшается в численномсоставеподдействиемсобытий,не имеющих отношения к обычным факторам естественного отбора.
Неслучайныескрещивания.Нарушения уравненияХарди-Вайнбергамогутпроисходить из-занеслучайныхскрещиваний.
Влияниеосновныхфакторовнаструктуру популяций описывается следующими формулами (Рис. 18.3).
Литература
Айала Ф., Кайгер Дж. Современная генетика, Т. 3. Москва, Мир, 72-166, 1988.
Алиханян С.И., Акифьев А.П., Чернин Л.С. Общая генетика. Москва, Высшая школа, 359-375, 1985.
Алтухов Ю.П. Генетика популяций и сохранение биоразнообразия. Соросовский образовательный журнал 1: 32-43, 1995.
Гершензон С.М. Основы современной генетики. Киев, Наукова Думка, 419-440, 1983.
Дубинин Н.П. Генетика. в кн.: Развитие биологии в СССР. Москва, Наука, 583-597, 1967.
Дубинин Н.П. Общая генетика. Москва, Наука, 334-372, 1970.
Дубинин Н.П., Глембоцкий Я.Л. Г е н е т и к а популяций и селекция. Москва, Наука, 1- 591, 1967.
Инге-Вечтомов С.Г. Генетика с основами селекции. Москва, Высшая школа, 454-475, 1989.
Кайданов Л.З. Генетика популяций. Москва, Высшая школа, 1-320, 1996.
Лобашов М.Е. Генетика. Издание второе. Ленинград, Издательство ЛГУ, 603-632, 1967.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяции. Москва, Наука, 1-277, 1973.
Четвериков С.С. О некоторых моментах эволюционного процесса с точки зрения современной генетики. Журнал эксперим. биологии. Сер. А, Т. 2, вып. 1: 3-54, 1926. Переиздано в книге “Классики советской генетики 1920-1940”, Ленинград, Наука, 133-170, 1968.
DobzhanskyT.G. Sergey Sergeewich Tshetverikov, 1880-1959. Genetics ##: 1-3, 1967.
RusselP.J.Genetics.Fifthedition.MenloParkCalifornia, AddisonWesleyLongman Inc.,714-771,1998.
405
Глава 18 |
|
|
Гены в популяциях |
|
|
|
|
|
|
|
|
|
|
|
Дополнение 18.1 |
|
|
|
|
Существование |
наследственной |
Однако, в начале 1900 годов наметилось |
||
изменчивости в природных популяциях |
противостояние |
дарвинизма |
è |
|
послужило исходным пунктом в цепи |
зарождающейся генетики. Согласно теориям |
|||
аргументов, приведенных Ч. Дарвином для |
Г. де Фриза и В. Иогансена, господствующее |
|||
доказательства того, что эволюция происходит |
прежде представление о всемогуществе |
|||
путем естественного отбора. По Дарвину, |
естественного отбора отвергалось и ему |
|||
некоторые из наследственных, изменений |
оставляли лишь сортирующую роль. К тому |
|||
обеспечивают их носителям больший успех в |
же факты существования наследственной |
|||
выживании и размножении по сравнению с |
изменчивости в популяциях диких животных |
|||
другими. В результате этого полезные |
и растений не были известны. |
|
||
изменения в ряду поколений будут |
Во второй половине 1920-x годов были |
|||
накапливаться, а вредные вытесняться. Это и |
сделаны первые шаги по объединению |
|||
называется процессом естественного отбора. |
Дарвиновской |
теории эволюции |
ñ |
|
|
|
|
|
|
Рисунок 18.3
à
II
отбор
p = p Wp-W W
I
p2 : 2pq : q2 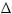 p = 0
p = 0
III
миграция  p = m(pm-p)
p = m(pm-p)
IV
дрейф
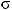 2q = 2Npq
2q = 2Npq
á
V
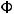 (p) = CW 2Np4Np-1q4Nmq-1
(p) = CW 2Np4Np-1q4Nmq-1
 p = 0
p = 0
Основные математические модели популяционной генетики: а) - панмиктическая популяция неограниченной численности, остающаяся в стабильном состоянии (F=0) в отсутствие отбора (W), миграции (M) и случайного дрейфа генов (N) - правило Харди-Вайнберга, представленное формулой I, где F è G - частоты генов, случайным образом комбинирующихся в гомозиготных (F , G ) и гетерозиготных (FG) генотипах. II, III, IV - элементарные формулы для отбора, миграции и случайного дрейфа генов как основных факторов, определяющих динамику частот генов в популяциях. б) - подразделенная популяция, представленная совокупностью субпопуляций, испытывающих одновременное воздействие случайного дрейфа и миграции генов, а также естественного отбора. V - математическая функция С. Райта для распределения частоты гена в такой популяции (Из: Алтухов, 1995).
406

Гены в популяциях |
|
|
|
Глава 18 |
|
|
|
|
|||
|
|
|
|||
принципами |
Менделевской генетики. |
генетическом конгрессе в Берлине. Они |
|||
Результат этого объединения известен как |
продемонстрировали насыщенность популяций |
||||
синтетическая теория эволюции. |
рецессивными мутациями. Начиная с 1930 года, |
||||
Первым исследователем, сделавшим шаг |
экспериментальными |
исследованиями |
|||
в направлении сближения дарвинизма и |
популяций занимается Н.П. Дубинин. |
||||
генетики был С.С. Четвериков (Рис. 1.17). В |
В 1927 году А.С. Серебровский |
||||
своей статье “О некоторых моментах |
предложил очень важный для генетики |
||||
эволюционного процесса с точки зрения |
популяций термин - “генофонд”, под которым |
||||
современной генетики”, опубликованной в 1926 |
понимал всю полноту генетических потенций |
||||
году, он дал единую концепцию эволюционного |
популяций, как природных, так и |
||||
процесса на |
основе менделирования, |
искусственных (породных и сортовых). |
|||
мутирования и отбора. Он показал, что |
Аналогичный термин (“gene pool”) в |
||||
свободное скрещивание (или панмиксия) в |
англоязычной литературе был предложен Ф.Г. |
||||
популяции служит механизмом стабилизации |
Добжанским в 1951 году. |
|
|
||
в ней частот генотипических классов |
В начале 1930-x годов на протяжении |
||||
гомозигот и гетерозигот. Установление такого |
нескольких лет появился ряд исследований, |
||||
равновесного соотношения частот генотипов, |
заложивших краеугольные камни в здание |
||||
по Четверикову, должно иметь следствием |
теории современной генетики популяций. В |
||||
накопление в популяции огромного фонда |
1930 году появилась работа Р.Фишера |
||||
скрытых мутаций. “...Вид, как губка, впитывает |
“Генетическая теория естественного отбора” |
||||
в себя гетерозиготные геновариации (мутации), |
(R.A. Fisher “The genetical theory of natural |
||||
сам оставаясь при этом внешне |
selection”), в 1931 году - работа Н.П. Дубинина |
||||
(фенотипически) однородным” - писал он. Тем |
“Генетико-автоматические процессы и их роль |
||||
самым снималось одно из существенных |
в процессах эволюции” и С Райта “Эволюция |
||||
возражений в отношении дарвинизма - тезис |
в менделевских популяциях” (S. Wright |
||||
о нехватке материала для действия отбора. С.С. |
“Evolution in mendelian populations”), â 1932 ãîäó |
||||
Четвериков обратил внимание на значение |
Дж. Холдейна “Факторы эволюции” (J.B.S. |
||||
колебаний численности популяций и на |
Haldane “Factors of evolution”) и Н.П. Дубинина |
||||
возникновение между ними изолирующих |
и Д.Д. Ромашова “Генетическое строение вида и |
||||
барьеров как на важные факторы эволюции. Он |
его эволюция”. В этих работах были заложены |
||||
особо подчеркивал активную роль отбора: |
основы экспериментального анализа проблемы |
||||
“...благодаря деятельности свободного |
генетики популяций. И, наконец, в 1937 году Ф.Г. |
||||
скрещивания и отбора в условиях |
Добжанский опубликовал книгу “Генетика и |
||||
менделевской наследственности, каждое даже |
происхождениевидов”(T.G.Dobzhansky “Genetics |
||||
слабое улучшение в организме имеет |
and the origin of species”), в которой, как многие |
||||
определенный шанс распространиться на всю |
считают, заложены основы современной |
||||
массу индивидов, составляющих свободно |
синтетической |
теории |
эволюции. В |
||
скрещивающееся сообщество (вид). |
дальнейшем, во всей литературе, кроме |
||||
...Дарвинизм - заключает Четвериков, - |
русскоязычной, только Ф. Добжанский, Р. |
||||
получил благодаря менделизму совершенно |
Фишер, С. Райт и Дж. Холдейнисталисчитаться |
||||
неожиданную и мощную поддержку”. |
основоположниками генетики популяций и |
||||
За год до этой публикации большая группа |
синтетической теории эволюции. Как будто и не |
||||
молодых исследователей (Б.Л. Астауров, Н.К. |
было вклада российских генетиков. |
|
|||
Беляев, С.М. Гершензон, П.Ф. Рокицкий, Д.Д. |
Вот, что пишет о С.С. Четверикове Ф.Г. |
||||
Ромашов) под руководством С.С Четверикова |
Добжанский |
(Dobzhansky, |
1967): |
||
выехала в экспедиции в южные районы |
“Плодотворные идеи часто восхитительно |
||||
Советского Союза для экспериментальной |
просты и не требуют многих страниц для их |
||||
проверки гипотезы о существовании мутаций в |
выражения. Грегор Мендель - основатель |
||||
природных популяциях дрозофилы. Уже в 1927 |
генетики. Он опубликовал только одну статью |
||||
году первые результаты этого анализа популяций |
в 1865 году, которая сделала его одним из |
||||
дрозофилы на Северном Кавказе были |
крупнейших ученых всех времен. Его другие |
||||
доложены Четвериковым на V международном |
статьи имеют не более чем биографический |
||||
|
|
|
|
|
|
|
|
|
|
|
|
407

Глава 18 |
|
|
Гены в популяциях |
|
|
|
|||
|
|
|||
интерес. Четвериков также опубликовал одну |
“Статья Четверикова 1926 года, хотя в ней |
|||
статью, которая позволила занять ему |
отсутствует математическая аккуратность, |
|||
почетное место в истории генетики. Название |
восхитительна по ясности рассуждений. В ней |
|||
Четвериковской классики в свободном |
доказывается (говоря современным языком), что |
|||
переводе на английский звучит как “On certain |
мутационный процесс является источником |
|||
aspects of the evolutionary process from the |
исходного материала для эволюции, а не самой |
|||
standpoint of genetics”. Она была опубликована |
эволюции.Прекрасновработепроанализированы |
|||
на русском языке во втором томе “Журнала |
следствия |
смешивания |
и неполного |
|
экспериментальной биологии (стр. 3-54) в |
наследования, т.е.возражения Дженкина Дарвину |
|||
1926 году. Короткое 2-x страничное резюме на |
против закона Харди (Четвериков был не знаком |
|||
английском языке мало помогло облегчению |
с вкладом Вайнберга). Сформулированы |
|||
восприятия этой статьи за пределами России. |
концепции генной частоты и генофонда (хотя и |
|||
Только в 1959 году отдельные пассажи его |
не даны этим понятиям названия). В ходе |
|||
статьи на английском языке были |
обсуждениягеографическойизоляциипопуляций, |
|||
опубликованы в качестве приложения к статье |
Четвериков близко подходит к понятию (хотя и не |
|||
Ф. Добжанского в издании трудов симпозиума |
описываетего)случайногогенетическогодрейфа. |
|||
“Gold Spring Harbor Symposia on Quantitative |
Естественный отбор по Четверикову является |
|||
Biology” (òîì 24). |
Полный перевод на |
ведущим |
фактором |
эволюционных |
английский, (перевел |
M.Barker, редактор M. |
преобразований, изменяющим частоты генов и |
||
Lerner) появился в 1961 году в Proceedings of the |
заменяющимодниаллелинадругие.Обсуждается |
|||
AmericanPhilosophicalSociety”(том105).Вместес |
значениеполиморфизма(анегетерозисакакодной |
|||
Р.А. Фишером, Сьюилом Райтом и Дж.Б.С. |
из причин отбора), подчеркивается важность |
|||
Холдейном, хотя и |
независимо от них, |
плейотропного действия гена. |
|
|
Четвериков четко, хотя и без элегантной |
Будучи экспериментатором и теоретиком, |
|||
математической утонченности, сформулировал |
Четвериков из своих теоретических рассуждений |
|||
основные принципы того, что позже стало |
сделал экспериментально |
проверяемые |
||
известно как биологическая, или синтетическая, |
предсказания.Онпредсказалсуществованиетого, |
|||
теория эволюции. Делались попытки на |
что сейчас известно как генетический груз. Имея |
|||
основании только даты публикации объявить в |
преимущество мудрости задним числом, нам |
|||
этом приоритет Четверикова. Сходные попытки |
сейчас трудно понять как некоторые биологи 40 |
|||
делались в пользу Фишера. Споры такого рода |
лет назад считали мутантов лабораторными |
|||
являютсяверхомтщетности.Всечетверопришли |
артефактами, которых невозможно найти в |
|||
к похожим идеям независимо, все четверо |
популяциях.Четвериковиегосотрудникирешили |
|||
являютсяосновоположниками,иэтопочетнодля |
найти мутантов в природе. На базе теоретических |
|||
всехчетверых.Четвериковнеимелвозможности |
соображений они разработали метод инбридинга |
|||
широко распространить свои идеи не только за |
и использовали его для анализа популяций |
|||
пределами, но даже и в самой России. Это не |
Drosophila melanogaster. Мутантов находили |
|||
было его виной, т.к. его научная деятельность |
весьма часто, большинство из них были |
|||
была грубо оборвана. Тем более было бы |
рецессивными, скрытыми в гетерозиготном |
|||
недобрым по отношению к нему, если бы мы |
состоянии”. |
|
|
|
попытались игнорировать его сейчас”. |
|
|
|
|
|
|
|
|
|
408