

Подготовка у универсиаде 2012 / Генетика (Жимулев) / 14ver7
.pdfГенетика определения пола |
Глава 14 |
|
|
Глава 14. Генетика определения пола
14.1. Общие принципы
Давно замечено, что соотношение мужского и женского полов в потомстве фактически от любого скрещивания близко соответствует соотношению 1:1, т.е. в достаточно большой группе потомков на 100 самцов рождается 100 самок. Совершенно очевидно,чтотакоерасщеплениекоррелирует с результатами особого генетического скрещивания-аналитического-когдаодиниз родителейгетерозиготен,другой-гомозиготен поанализируемомупризнаку.Поэтомуapriori можнопредположить,чтоодинизполовкакбы гетерозиготен,адругойгомозиготенпофактору, определяющемупол.Цитологическийанализ выявляетнаиболеезаметноеразличиеуполовсочетание половых хромосом в кариотипе, причемодинполимеетодинаковыехромосомы, в кариотипе другого - две разных половых хромосомы,т.е.этотполкакбы“гетерозиготен”.
Привлекает внимание огромное разнообразиевариантовполовогоразмножения и соответствующих ему версий определения пола,затрудненадажеихклассификация.
Выделяютнесколькотиповопределения полавзависимостиотчислаисоставаполовых хромосом, например, у самца могут быть X- и Y-хромосомы, а у самки - XX. К такому типу относятся человек, дрозофила, водяной клоп Ligaeus,поименикоторогоиназванданныйтип определенияпола,атакжемногиедругиевиды животных.
Ещеодинтип,названныйпоименидругого водяного клопа Protenor, и встречающийся у некоторыхбабочекичервей,связансналичием у самцов X0 хромосом, а у самок двух X- хромосом.
Другой тип хромосомного определения пола найден у птиц, некоторых бабочек, рыб, земноводных и цветковых растений. У них гетерогаметным (т.е. с разными половыми хромосомами)поломявляетсяженский,исамки имеютнаборполовыххромосом ZW или Z0,в товремякаксамцы-ZZ.
Известны и некоторые другие типы определенияпола.
Совершенно очевидно, что в перечисленных выше случаях колоссальные
èлегкофиксируемыеразличияворганизации
èфизиологии особей разных полов развиваются на фоне идентичных у самцов и самок наборов генов, расположенных в большей части хромосом, не связанных с половыми различиями кариотипов, - аутосомах.Инапротив,всеполовыеразличия определяются сравнительно небольшим числом генов, заключенных в половых хромосомах.
Возникает вопрос, каким образом варьирование числа и состава половых хромосом может определять развитие мужского или женского полов? Еще более интересно, что, например, у дрозофилы, в Y- хромосоме генов практически нет (к настоящемувремениоткрытотолько11генов, влияющихнаформированиесперматозоидов - см. Раздел 9.5.10.). Более того, при потере Y-хромосомы, особь X0 также является самцом. А особь XXY - это нормальная плодовитая самка. Каким же образом формируются самцы и самки у дрозофилы?
Различают дифференциацию пола (фенотипический пол), то есть появление внешних гениталиев, вторичных половых признаков, и первичное определение пола. Подпервичнымопределениемполапонимают появление гонады (репродуктивного органа соматической природы) самки или самца - яичника или тестиса. Считается, что принципиальная схема этого процесса консервативна. Существует конкретный контролирующийсигнал,включающийнекий ключевой ген. Этот ген, в свою очередь, активирует некое число детерминант гонадогенезиса и далее факторов дифференцировки половых признаков. Все компоненты этой системы могут различаться у разных животных. В Табл. 14.1 представлены принципиальные схемы определенияполаунекоторыхпредставителей животного мира, обозначены соответствующие гены.
337

Глава 14 |
Генетика определения пола |
|
|
Таблица 14.1
Принципиальная схема определения пола у некоторых видов животных и растений (Из: Смирнов, 1997, стр. 27).
|
Saccharomyces |
Caenorhabditis |
Drosophila |
Alligator |
Mammalia |
|
cerevisiae |
elegans |
melanogaster |
mississippiensis |
|
|
|
|
|
|
|
Контролирующий |
Транскрипция |
Соотношение Х-хромосом |
Внешняя |
Y-хромосома |
|
сигнал |
ÍÎ-ãåíà |
и аутосом |
|
температура |
|
|
|
|
|
|
|
Ключевой ген |
ÌÀÒ (α /a) |
her (+/-) |
Sxl (+/-) |
TDF |
TDF-SRY |
|
|
|
|
|
|
Ãåíû, |
- |
xoll, sdc1, sdc2, |
sis-a, sis-b, |
Эффекторные |
Гормоны |
контролирующие |
|
herl, tra2, tra3, |
da, liz, fl(2)d, |
молекулы, |
|
гонадогенез |
|
ferm1, ferm2, |
Sxl, tra, |
гормоны |
|
(определяющие |
|
ferm3, tra1 |
tra2, dsx, ix |
|
|
ïîë) |
|
|
|
|
|
|
|
|
|
|
|
Ãåíû, |
+/- |
+/- |
+/- |
+/- |
+/- |
контролирующие |
|
|
|
|
|
половую |
|
|
|
|
|
дифференцировку |
|
|
|
|
|
Примечание. + и - означают альтернативу включения или выключения соответствующего гена.
В некоторых случаях появление мужского или женского пола определяется ненаследственнымиразличиями,авозникает под влиянием условий среды. Классическим примером служит морской червь Bonellia viridis. Самцы размером в несколько миллиметров живут в матке самки, где выполняют свою задачу - оплодотворяют яйцеклетки. Самец является типичным паразитом, живущим внутри тела самки, размер которой примерно равен размеру сливы (Рис. 14.1).
Свободно плавающие личинки, развивающиеся после оплодотворения яйцеклеток, некоторое время ведут свободный образ жизни, а затем прикрепляются к хоботу половозрелой самки,либооседаютиприкрепляютсякодну. Личинки этих двух сортов ничем друг от друга не отличаются. Куда попадает данная личинка - это дело случая. Однако во что превращается личинка, т.е. будет ли она самцомилисамкой, определяется условиями в период несвободной жизни личинки. Личинки, прикрепившиеся к хоботу самки, развиваются в самцов. Они проникают в женские половые органы и живут там как паразиты. Личинки, прикрепившиеся ко дну, развиваются в самок.
Рисунок 14.1
Самка
Размер
самца
Самец (сильно увеличено)
Половые различия между самкой и самцом у морского червя Bonellia viridis (Из: Дубинин, 1970, стр. 121).
Определение пола у миссисипских крокодилов регулируется изменением внешней температуры (Табл. 14.1, см. более подробно у Pieau, 1996).
Литература к разделу 14.1.
Гершензон И. Основы современной генетики. Киев, Наукова Думка, 101-135, 1983 Гершкович И. Генетика. Москва, Наука, 110-
124, 1968.
Дубинин Н.П. Общая генетика. Москва, Наука, 386-394, 1970.
Лобашев М.Е. Генетика, Ленинград, изд-во ЛГУ, 202-225, 1967.
338

Генетика определения пола |
Глава 14 |
|
|
Смирнов А.Ф. Молекулярно-генетические |
|
Рисунок 14.2 |
|
механизмы первичной детерминации |
|
пола у млекопитающих. Соросовский |
|
образовательный журнал 1: 26-34, 1997. |
|
Pieau C. Temperature variation and sex determination |
|
in reptiles. BioEssays 18: 19-26, 1996. |
|
14.2. Гинандроморфы, интерсексы, гермафродиты
èдругие половые отклонения
Óдрозофилы и у других организмов известны случаи гинандроморфизма, когда разные участки тела по своим признакам принадлежат разным полам. Организм выглядит как мозаик, у которого одна часть мужская, а другая - женская (Рис. 14.2). В данном случае зигота имеет две Х- хромосомы и должна бы развиться в самку.
Она является гетерозиготой по генам |
|
Билатеральный гинандроморф у Drosophila |
|||
белоглазия и маленьких крыльев w m/w+m+, |
|
melanogaster (из: Дубинин, 1970, стр. 386). |
|||
расположенных в X-хромомсоме. Во время |
интерсексы отличаются от нормальных |
||||
|
|
+ + |
|||
первых делений дробления хромосома w m |
особейтем,чтоунихпервичныеивторичные |
||||
утрачивается и если экватор митотического |
|||||
половые признаки носят промежуточный |
|||||
деления располагается по линии симметрии |
характер, образуя непрерывный ряд |
||||
от головной до хвостовой части эмбриона, |
переходов от нормального самца к |
||||
одна половина (левая) тела мухи состоит из |
нормальной самке. |
||||
клеток, имеющих только одну Х-хромосому, |
|
Наряду с разнополостью у многих |
|||
что соответствует генотипу самца. Правая |
|
||||
растений и у низших животных мужской и |
|||||
сторона имеет две Х-хромосомы и |
женский пол совмещается в одном |
||||
развивается в самку. |
|
|
организме, который таким образом является |
||
У непарного шелкопряда Lymantria |
|||||
гермафродитом. |
|||||
dispar имеются резкие различия между |
|
|
|||
самками и самцами. Скрещивания разных |
Литература к разделу 14.2. |
||||
географических |
ðàñ |
этой бабочки |
Гершкович И. Генетика. Москва, Наука, 110- |
||
(европейских и японских) привело к |
124, 1968. |
||||
появлению форм, переходных по своим |
|||||
Дубинин Н.П. Общая генетика. Москва, Наука, |
|||||
признакам между самцами и самками, т.е. к |
386-394, 1970. |
||||
появлениюинтерсексуальности. Интерсексы |
Allen G.E. Thomas Hunt Morgan: the man and his |
||||
обнаружены и у дрозофилы. |
|
science, Princeton, New Jersey, Princeton |
|||
От гинандроморфов интерсексы |
|
University Press, 241-249, 1978. |
|||
отличаются тем, что у них отсутствуют |
|
|
|||
различно детерминированные по полу |
14.3. Балансовая теория |
||||
сектора (Рис. 14.3). |
|
|
определения пола у дрозофилы |
||
У интерсексов до определенного |
В 1921 году один из основоположников |
||||
|
|
|
|||
момента развития сохраняется генетически |
современной генетики Кальвин Бриджес |
||||
детерминированный пол, но затем развитие |
обнаружил несколько самок, имевших |
||||
продолжается |
â |
направлении |
триплоидный набор хромосом, т.е. 3X+3A-три |
||
противоположного пола. В результате |
набора X-хромосом и три набора аутосом. В |
||||
339

Глава 14 |
|
Генетика определения пола |
|
|
|
|
|
|
Рисунок 14.3 |
|
|
à |
|
á |
â |
ã |
ä |
Самец (а), самка (б) и некоторые ненормальные половые типы дрозофилы: интерсекс (в), сверхсамка (г), сверхсамец (д) (Из: Гершкович, 1968, стр. 112).
результате скрещивания этих самок с нормальными самцами (2A+XY) в потомстве среди нормальных самок и самцов были обнаружены особи с промежуточным или необычным проявлением половых признаков. Все потомство распалось на 8 классов в зависимости от соотношения половых хромосом и аутосом:
1.3X:3A - триплоидная самка
2.2X:2A - диплоидная самка
3.(2X+Y):2A - самка.
Вэтихтрехслучаяхсоотношениечисла X-хромосом к числу аутосом составляет единицу. Наличие мужской Y-хромосомы не влияет на нормальное развитие самки.
4. Особи, имеющие хромосомную конституциюXY:2A,т.е.укоторыхотношение числа X-хромосом к числу аутосом составляет 0.5, были нормальными самцами.
5 и 6. Интересными оказались особи 2X:3A и (2X+Y):3A, у которых отношение числа X-хромосом к числу аутосом было промежуточным между 0.5 и 1. Они имели смешанное проявление мужских и женских половых признаков. Такие особи были интерсексами (см. Рис. 14.3).
7 и 8. И, наконец, если число наборов аутосом увеличивалось до трех при наличии одной X-хромосомы (X:3A), развивался
“сверхсамец” - организм с гипертрофированными признаками самца, однако стерильный, слабый и быстро погибающий.
Напротив, увеличение числа X- хромосом при диплоидном наборе аутосом (3X:2A) ведет к формированию “сверхсамки” с ненормально развитыми яичникамиидругиминарушениямипризнаков пола. Они слабые и быстро погибают.
К. Бриджес пришел к выводу, что не присутствие двух X-хромосом определяет женский пол и не наличие Y-хромосомы определяет мужской пол у дрозофилы. Пол, по его мнению, определяется балансом числа X-хромосом и наборов аутосом, т.е. Y- хромосома у дрозофилы вообще не играет роли в определении пола.
Литература к разделу 14.3.
Allen G.E. Thomas Hunt Morgan: the man and his science, Princeton, New Jersey, Princeton
University Press, 241-249, 1978.
14.4. Действие генов при определении пола у дрозофилы
Каким образом соотношение числа X- хромосом и аутосом, согласно балансовой теории Бриджеса, может влиять на развитие
340
Генетика определения пола |
|
|
Глава 14 |
|
|
|
|
|
|
|
|
Дополнение 14.1 |
|
|
|
В то время как К. Бриджес считал идею |
Морган не мог принять заключение Бриджеса |
||
балансового определения пола неотразимой, |
о том, что большинство генов, необходимых |
||
Т. Морган никогда ее полностью не одобрял. |
для развития самца, были в аутосомах, а |
||
В письме Отто Мору в 1923 году Морган ясно |
множество |
генов, |
определяющих |
выразил свой скептицизм: “Я полностью |
формирование самок, расположено в X- |
||
согласен с критикой балансовой теории, |
хромосоме. Он писал: “То, что следует из |
||
которую Бриджес так неумеренно любит. Она |
данных на триплоидах, не дает какой-либо |
||
есть и будет оставаться выдумкой до тех пор, |
особой информации о наличии генов для |
||
пока мы не сможем привлечь хоть какие-то |
определения пола. Если мы думаем о |
||
объективные критерии для ее оценки. Она |
хромосомах, только как особых вместилищах |
||
может оказаться, и, возможно, является |
генов, из этого следует, что такие гены |
||
истинной, однако она говорит нам не больше, |
существуют, но нет никаких свидетельств того, |
||
чем сами полученные факты. В этом я убеждал |
какие они. Если даже гены [в этот процесс] |
||
Бриджеса с самого начала, но безрезультатно”. |
вовлечены, мы не можем судить о том один |
||
В своей книге “ Теория гена” в 1926 году |
ли ген, который влияет на развитие самки, |
||
Т. Морган объяснил идею Бриджеса очень |
расположен в X-хромосоме, или там сотни |
||
коротко, на двух страницах, без какого-либо |
таких генов. То же самое можно сказать и о |
||
одобрения ее. Его возражения свелись к двум |
генах во всех хромосомах, определяющих |
||
пунктам: во-первых, балансовая теория только |
признаки самца. То ли они есть во всех |
||
пересказывала факты, т.е. перефразировала |
хромосомах или только в одной паре |
||
экспериментальные данные о корреляции |
хромосом”. |
Морган |
также возражал |
между половыми фенотипами и отношениями |
цифровым аспектам теории Бриджеса (Из: |
||
чисел Х-хромосом и аутосом. Во-вторых, |
Allen, 1978, pp. 245-246). |
|
|
пола? В последние годы у дрозофилы |
в свою очередь, контролирует 3 различных |
||
открыты многочисленные гены, влияющие |
направления дифференцировки пола: |
||
направильноеформированиепола,срединих |
формирование половых признаков в |
||
такие, как Sxl (Sex lethal), da (daughterless), |
соматических клетках и в клетках |
||
sis (sisterless), tra (transformer), dsx (double |
зародышевого пути, а также контроль |
||
sex) è äð. |
дозовой компенсации (Рис. 14.4). Каким |
||
Эксперименты показывают, что |
образом это происходит? |
||
результаты соотношения числа X-хромосом |
На начальных этапах формирования |
||
и аутосом улавливаются геном Sxl на ранней |
пола у эмбрионов действуют гены sis-a и sis- |
||
стадии эмбрионального развития. Этот ген, |
b, расположенные в X-хромосоме, и ген da, |
||
Рисунок 14.4
 дозовая компенсация
дозовая компенсация
X:A |
|
Sxl |
половые признаки |
|
|
|
в соматических тканях |
||
|
|
|||
|
|
|
|
|
половые признаки в зародышевых клетках
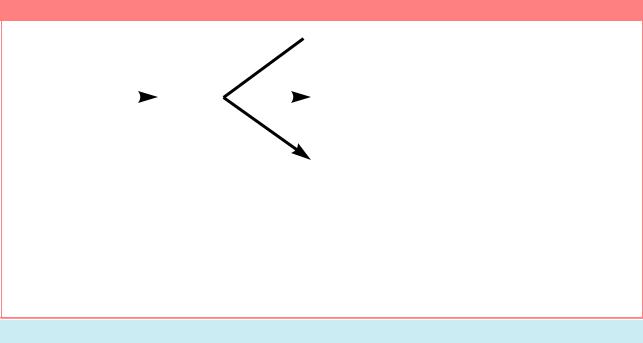
Первичный сигнал, возникающий из соотношения числа Х-хромосом и аутосом, контролирует все аспекты половой дифференцировки через действие ключевого гена Sxl. У самок, имеющих отношение X:A, равное единице, Sxl активен. У самцов, имеющих отношение X:A, равное 0.5, ген Sxl остается неактивным. Состояние активности гена Sxl регулирует развитие трех процессов, находящихсяподегоконтролем:дозовойкомпенсации,развитияполовыхпризнаковвсоматических и зародышевых клетках (Из: Nö thiger, 1992, ð. 177).
341
Глава 14 |
Генетика определения пола |
|
|
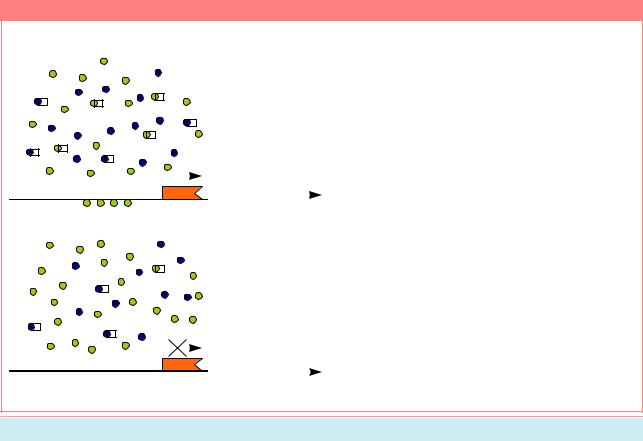
расположенный в аутосоме. Белковые продукты этих генов образуют комплексную молекулу белка. Продукт гена da поступает в яйцеклетку из организма матери, его количество всегда соответствует двум дозам, т.к. он считывается с генов, локализованных в двух материнских аутосомах. Количество продуктов, считанных с генов sis-a и sis-b, зависит от того, сколько X-хромосом у эмбриона - две или одна. В клетках эмбриона женского пола две Х-хромосомы. С них считываются продукты генов sis-a и sis-b в двух дозах. У самцов только одна Х-хромосома, которая в условиях нефункционирующего механизма дозовой компенсации на ранних стадиях развития производит только одну дозу продуктов этих генов. Поэтому комплекс белков sis/da имеет отношение составляющих его компонентов 1:2 у самцов или 1:1 у самок. Эти белковые продукты поступают на регуляторную зону ключевого гена, определяющего пол - Sxl. Эта зона содержит 2 участка, стимулирующих транскрипцию РНК с данного гена: ранний и поздний промоторы (PE è PL) (Рис. 14.5). Только в том случае если комплексный белок sis/da содержит 2 дозы sis, он может
активироватьначалотранскрипциисраннего промотора (PE на Рис. 14.5). Это происходит в самом раннем эмбриональном развитии, на стадии бластодермы. Позднее транскрипция может начаться и с позднего промотора как у XX/AA, так и X/AA особей. Но результаты включения транскрипции с каждого из этих промоторов будут разными. Рассмотрим их (Рис. 14.6).
Ген Sxl содержит 8 участков, кодирующих последовательность аминокислот (1-8 на Рис. 14.6) - экзонов, разделенныхнекодирующимирайонами(Рис. 14.6). У самцов (X:A=0.5) при активировании позднего промотора (PL) считывается третий экзон (Рис. 14.6A), содержащий большое число кодонов UGA, после каждого из которыхтрансляцияостанавливается,ибелок получается усеченным. В отсутствие нормального функционального белка гена Sxl, ген tra, расположенный далее в данном каскаде, также функционирует неправильно, опятьжедаваякороткуюнефункциональную молекулу белка (трансляцию блокирует кодон UGA во втором экзоне - см. Рис. 14.6A). Хотя белок другого гена - tra2 (см. Рис. 14.6) - присутствует у обоих полов, он не формируется до нормального состояния
Рисунок 14.5
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Sxl |
|
|
PL |
|
|
|
PE |
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
PL |
|
PE |
Sxl |
|
|
|
Модель механизма включения гена Sxl |
||
À |
|
|
у разных полов. У самок (А) и самцов |
||
|
|
|
(Б), белки, кодируемые генами из X- |
||
|
|
|
хромосомы |
(прямоугольники) |
|
|
|
|
формируют активные комплексы с |
||
|
|
|
белками, |
кодируемыми генами, |
|
|
|
|
расположеннымиваутосомах(кружки). |
||
|
|
|
Самки имеют в два раза больше |
||
Sxl ON |
комплексных молекул, чем самцы. |
||||
|
|
Самка |
Наличие большого числа этих молекул |
||
Á |
|
|
позволяет им связываться с PE |
||
|
|
|
промоторным элементом гена Sxl и |
||
|
|
|
активировать транскрипцию (стрелка |
||
|
|
|
вправо - SxlON). У самцов (Б) число |
||
|
|
|
молекул |
комплекса невелико и |
|
|
|
|
неспособно активировать PE промотор. |
||
|
|
|
Ген Sxl не включается (SxlOFF), ÷òî è |
||
Sxl |
OFF |
приводит в конечном счете к развитию |
|
|
|
||
|
|
Самец половых признаков самца (Из: Belote, |
|
|
|
|
|
|
|
|
1992, p. 324). |
342

Генетика определения пола |
|
Глава 14 |
|
|
|
|
|
|
Рисунок 14.6 |
|
|
À |
Á |
|
X:A = 0.5 |
X:A = 1.0 |
|
AUG UGA |
AUG |
UGA |
Sxl |
|
|
|
|
|
|
|
1 |
2 |
3 |
4 |
5 6 7 |
8 |
1 |
2 |
нефункциональный белок Sxl
AUG UGA |
AUG |
tra |
|
|
|
1 |
2 |
3 |
1 |
нефункциональный tra2 белок tra
3 |
4 |
5 6 7 |
8 |
|
Sxl |
|
|
|
|
UGA |
|
|
2 |
3 |
|
|
tra |
|
|
AUG |
UGA |
AUG |
UGA |
dsx |
|
|
1 |
2 |
3 |
5 |
6 |
1 |
2 |
3 |
4 |
ix
dsxM |
dsxF |
подавление дифференцировки |
подавление дифференцировки |
самки |
самца |
Самец |
Самка |
Каскад генных взаимодействий, приводящих к формированию соматических половых признаков самца (А) и самки (Б) (Из: Belote, 1992, p. 327). В каскад вовлечены гены Sxl, tra, tra2, ix и dsx. Прямоугольники с цифрами обозначают кодирующие части генов - экзоны. Зачерненные части экзонов представляют собой участки, кодирующие аминокислоты. Зигзагами показаны интроны.
вотсутствиефункциональногопродуктагена tra. Более того, в отсутствие нормальных продуктов генов tra и tra2, формируется особый dsxM (самцовый) белок, считанный с определенного набора экзонов (см. Рис. 14.6A). Такой белок репрессирует развитие признаков женского пола. В результате нарушений в каскаде этих генов, развитие направляется в сторону формирования признаковсамца.
У самок (X:A=1) транскрипт гена Sxl не содержит экзона номер 3 со стоп-кодоном, в результате чего формируется полноценный белок Sxl, взаимодействующий с геном tra, который в комплексе с белком гена tra2 регулирует образование специфической для самок РНК гена dsxF (см. Рис. 14.6Б). Наличие специфического для самок продукта dsxF способствует вовлечению в данный каскад гена ix. Белки генов dsxF è ix
343
Глава 14 |
Генетика определения пола |
|
|
инактивируют многие гены, которые могли бы привести к формированию самцов, и в конечном итоге формируется самка.
По этой схеме формируются внешние (соматические) половые признаки.
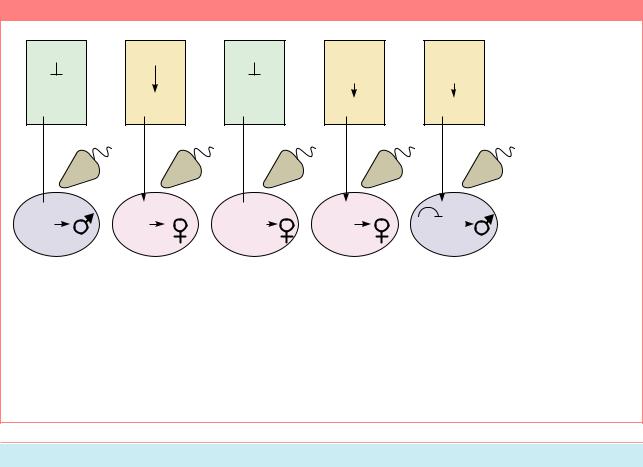
Под четким генетическим контролем находится определение пола у комнатной мухи Musca domestica. Y-хромосома самца обычно несет фактор, определяющий пол самца - M;, которыйи составляет первичный сигналопределенияпола.Число X-хромосом на определение пола не влияет. У представителей других популяций этот фактор локализуется в одной из аутосом и даже в X-хромосоме. Это предполагает, что он является мобильным элементом. В линиях с аутосомной локализацией M- фактора Y-хромосома может быть утрачена, все особи несут только X-хромосомы, но при этом формируются самцы.
И,наконец,втретьемтипелинийсамцы и самки являются гомозиготами по аутосомному M-фактору. Каким же образом в линии с M/ M у обоих полов формируются самки? Оказывается, это достигается действием доминантного аллеля гена F, расположенного в 4-й хромосоме. Аллель F,
оказывает эпистатическое действие на ген M и определяет развитие самки в присутствие одной или нескольких копий M-фактора. Нормальный аллель - F+ негативно контролируется M-факторами. Таким образом доминантный ген FD стимулирует развитие самок, рецессивный F+ - развитие самца. Современные представления о генетическомконтролеполаукомнатныхмух изображены на Рис. 14.7.
Интересно, что последовательность ДНК, гомологичная гену Sxl Drosophila melanogaster, присутствует в геноме Musca domestica, однако он экспрессируется как у самок, так и самцов: в ходе развития обнаружены те же самые транскрипты и изоформы белков у обоих полов.
У средиземноморской плодовой мушки Ceratitis capitata также присутствует ген, гомологичный гену Sxl Drosophila melanogaster. Однако, и у этого вида данный ген экспрессируется у обоих полов.
Очевидно, что первичные сигналы, активирующие каскады генной активности, приводящие к развитию разных полов, сильно различаются у разных видов.
Рисунок 14.7 |
|
|
|
|
|
À |
Á |
 |
à |
Ä |
|
M |
M;FD(ON) |
M |
íåò M |
íåò M |
|
F(OFF) |
|
F(OFF) |
F(ON) |
F(ON) |
яйцеклетка |
(íåò |
продукт F |
(íåò |
продукт F |
продукт F |
|
продукта F) |
продукта F) |
|
|||
|
|
|
|
||
F+ |
F+ |
M;FD |
F+ |
M;F+ |
сперматозоид |
(íåò Ì) |
(íåò Ì) |
|
(íåò Ì) |
|
|
F(OFF) |
F(ON) |
M;FD(ON) |
F(ON) |
M;F(OFF) |
зигота |
Схемы, объясняющие формирование самцов и самок у Musca domestica (Из: Dü bendorfer, Hediger, 1998). A – активность гена M отключает ген F при созревании яйца. В отсутствие продукта гена F в яйцеклетке формируется самец. Б – активность гена FD подавляет действие гена M, в результате чего в яйцеклетке появляется продукт гена F; развивается самка. В – в яйцеклетке нет продукта гена F, но в зиготу вносится ген FD сперматозоидом; развивается самка. Г – в отсутствие гена M в яйцеклетке и сперматозоиде для формирования самки достаточно гена F+. Д – в отсутствие FD в яйцеклетке и сперматозоиде, наличия одной дозы M в сперматозоиде достаточно для формирования самца.
344

Генетика определения пола Глава 14
Литература к разделу 14.4. |
|
При наличии трех или даже четырех X- |
|||||||
|
хромосом, но в присутствии Y-хромосомы |
||||||||
Корочкин Л.И. Введение в генетику развития. |
|||||||||
формируется мужской тип тела, правда тоже |
|||||||||
Москва, Наука, 210-229, 1999. |
|
с отклонениями, известными под названием |
|||||||
Belote J.M. Sex determination in Drosophila |
|||||||||
melanogaster: from the X:A ratio to |
“синдрома Клайнфелтера” (см. Дополнение |
||||||||
doublesex. Seminars in Devel. Biol. 3: 319- |
4.1.). |
|
|
|
|
||||
330, 1992. |
|
|
Уже в самых первых актах |
||||||
Cline T.W., Meyer B.J. Vive la difference: males vs |
детерминации |
гонад |
ó |
самцов |
|||||
females in flies vs worms. Annual Rev. Genet. |
млекопитающих необходима активность Y- |
||||||||
30: 637-702, 1996. |
|
|
хромосомы, точнее сцепленного с ней |
||||||
Dübendorfer A., Hediger M. The female determining |
тестис-определяющего фактора (testis- |
||||||||
gene F of the housefly, Musca domestica, acts |
|||||||||
determining factor - TDF). Этот фактор был |
|||||||||
maternally to regulate its own zygotic activity. |
|||||||||
идентифицирован в результате анализа |
|||||||||
Genetics 150: 221-226, 1998. |
|
|
транслокаций небольших фрагментов Y- |
||||||
Graves J.A.M. The origin and function of the |
|||||||||
mammalian Y chromosome and Y-borne genes |
хромосомы на Х-хромосому, в результате |
||||||||
- an evolving understanding. BioEssays 17: |
чего формировались особи с инверсией пола |
||||||||
311-321, 1995. |
|
|
- самцы ХХ и самки XY. Такие отклонения |
||||||
|
|
встречались у отдельных особей - |
|||||||
Hediger M. et al., The male-determining activity on |
|||||||||
the Y chromosome of the house fly (Musca |
представителей |
17 |
|
семейств. |
|||||
domestica L.) consists of separable elements. |
Инвертированные самцы мышей XXSxr â Õ- |
||||||||
Genetics 150: 651-661, 1998. |
|
|
хромосомах имеют перенес¸нные из Y- |
||||||
Meise et.al., Sex-lethal, the master sex-determining |
хромосомы небольшие фрагменты. Они |
||||||||
gene in Drosophila, is not sex-specifically |
|
обладают тестисами, но их репродуктивная |
|||||||
regulated in Musca domestica. |
|
|
|||||||
|
|
система дефектна из-за |
аномалий |
||||||
Development, V. 125, P. 1487-1494, 1998. |
|||||||||
сперматогенеза. |
|
|
|
||||||
Nö thiger R. Genetic control of sexual development |
|
|
|
||||||
В 1990 году было установлено, что у |
|||||||||
in Drosophila. Verh. Dtsch. Zool. Ges. 85: |
|||||||||
человека фактор TDF это не что иное как ген |
|||||||||
177-183, 1992. |
|
|
SRY (Sex determining Region Y gene), |
||||||
Saccone G. et al., The Ceratitis capitata homologue |
|||||||||
который располагается в коротком плече Y- |
|||||||||
of the Drosophila sex-determining gene Sex- |
|||||||||
lethal is structurally conserved, but nof sex- |
хромосомы во фрагменте длиной 35 т.п.н. |
||||||||
specifically regulated. Development 125: |
Ген SRY содержит консервативный |
||||||||
1495-1500, 1998. |
|
|
домен (HMG), кодирующий фрагмент |
||||||
14.5. Определение пола |
|
|
белковой |
молекулы |
размером 80 |
||||
|
|
аминокислотных остатков. Белковый |
|||||||
у млекопитающих |
|
|
комплекс, кодируемый доменом HMG, |
||||||
Ïðè |
определении |
ïîëà |
ó |
специфически связывается с ДНК, приводя |
|||||
млекопитающих Y-хромосома играет |
к изгибанию е¸ молекулы. Такая деформация |
||||||||
решающую роль. Известно, что XX и XY - |
структуры ДНК, индуцируемая SRY-белком |
||||||||
соотношения половых хромосом приводят |
илиродственнымиемумолекулами(известно |
||||||||
к образованию нормальных женской и |
более 100 белков с HMG-доменом), может |
||||||||
мужской особей, соответственно. Однако, |
механически передаваться на достаточно |
||||||||
при отсутствии Y-хромосомы формируется |
большое расстояние и играть важную роль в |
||||||||
особь женского пола при любом числе X- |
регуляциитранскрипции,репликациии |
||||||||
хромосом. Особь X0 развивается главным |
рекомбинации. |
|
|
|
|||||
образом по женскому типу, но имеет ряд |
Только HMG-домен является |
||||||||
отличий от нормы, известных под названием |
консервативной частью гена SRY. За |
||||||||
“синдром Тернера” (см. Дополнение 4.1.). |
|
пределами |
ýòîé |
последовательности |
|||||
|
|
|
|
обнаружены существенные различия между |
|||||
345
Глава 14 |
|
|
|
|
|
|
|
Генетика определения пола |
|||||
гомологичными генами даже у Рисунок 14.8 |
|
|
|
|
|
|
|||||||
близкихвидов.Например,учеловека |
Гермафродит XX |
|
|
|
|
||||||||
этот ген имеет небольшой размер, не |
|
|
|
|
|||||||||
|
|
|
TRA-2A |
|
|
|
|
||||||
содержитинтроновикодируетбелок |
|
|
|
|
|
|
|
|
|||||
размером в 204 аминокислотных |
|
|
|
|
|
TRA-2A |
|
|
|||||
|
|
|
|
|
|
|
|
||||||
остатка. Его гомолог, выделенный из |
|
|
|
FEMs |
FEMs |
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
генома мыши, содержит уже 395 |
|
|
|
|
|
|
|
|
|||||
аминокислот. |
|
|
|
|
|
|
|
|
|
|
|
||
Кроме гена SRY Y-хромосома |
|
|
|
|
|
|
|
|
|||||
содержит |
несколько |
генов, |
|
|
|
TRA-1 |
|
|
|
|
|||
необходимых для сперматогенеза. |
|
|
|
|
her-1 |
|
|
|
|
||||
Литература к разделу |
|
|
ÿäðî |
|
|
|
|
|
|||||
|
|
|
sdc-1 |
|
|
fem-1 |
|
|
|||||
14.5. |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
X:A |
xol-1 |
sdc-2 |
her-1 |
tra-2 |
fem-2 |
tra-1 |
|
|
|
|
|
|
|
|
1.0 |
|
sdc-3 |
|
|
fem-3 |
|
|
Корочкин Л.И. Введение в генетику |
|
|
|
|
laf-1 |
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
развития. Москва, Наука, 210- |
|
|
|
|
|
|
|
|
|||||
229, 1999. |
|
|
|
Самец X0 |
|
|
tra-3 |
|
|
|
|||
Смирнов |
À.Ô. |
Молекулярно- |
|
-1 |
|
|
|
|
|||||
|
|
|
|
|
|
|
|||||||
генетические |
механизмы |
|
|
|
HER |
|
|
|
|
||||
|
|
|
|
|
|
|
|
||||||
первичной детерминации пола |
|
|
|
TRA-2A |
|
|
|
|
|||||
у млекопитающих. Соросовский |
|
|
|
|
HER |
|
|
||||||
|
|
|
|
|
1 |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
- |
|
|
образовательный журнал N1, |
|
|
|
FEM-2 TRA-2A |
|
|
|||||||
26-34, 1997. |
|
|
|
|
|
|
FEM-1 |
FEM-3 |
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
HER-1 |
|
14.6. Определение пола |
|
|
|
|
TRA-1 |
|
|
|
|
||||
у нематоды |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
her-1 |
HER-1 |
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
||
Caenorhabditis elegans |
|
|
|
ÿäðî |
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
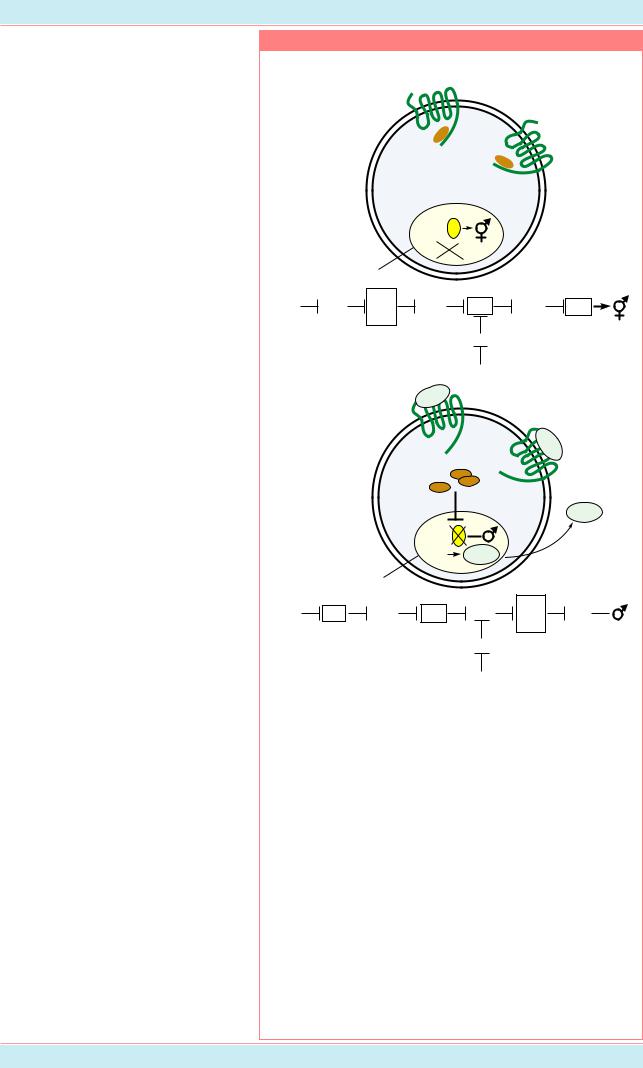
У этого вида нематоды два |
|
|
sdc-1 |
|
|
fem-1 |
|
|
|||||
пола: самцы X0 и гермафродиты XX. |
X:A |
xol-1 |
sdc-2 |
her-1 |
tra-2 |
fem-2 |
tra-1 |
|
|||||
0.5 |
|
sdc-3 |
|
|
fem-3 |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|||
Гермафродиты в данном случае - это |
|
|
|
|
laf-1 |
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
самофертильные самки, у которых |
|
|
|
|
|
|
|
|
|||||
признак самцовости заключается |
|
|
|
|
tra-3 |
|
|
|
|||||
лишь в образовании ограниченного |
Иерархия взаимодействия генов, которые контролируют |
||||||||||||
числа |
|
сперматозоидов, |
определение соматических половых признаков и |
||||||||||
используемых исключительно для |
дифференцировку у гермафродитов и самцов. На каждом |
||||||||||||
внутреннего самооплодотворения. |
|
рисунке в нижней части показаны каскады генов, в |
|||||||||||
Самцыобразуютсяврезультате |
верхней – модели молекулярных механизмов. Гены в |
||||||||||||
регуляторном каскаде, которые функционально активны, |
|||||||||||||
очень |
редкого |
спонтанного |
заключены в прямоугольники и набраны жирном |
||||||||||
нерасхождения X-хромосом во |
шрифтом. Темная полоса указывает на наличие |
||||||||||||
время размножения гермафродитов |
негативного регуляторного влияния. Линии, набранные |
||||||||||||
или в результате расщепления от |
пунктиром, указывают на отсутствие негативного |
||||||||||||
скрещивания |
самцов |
ñ |
действия, поскольку выше расположенный регулятор |
||||||||||
гермафродитами. Пол определяется |
неактивен. Наиболее вероятно, что прямой мишенью |
||||||||||||
балансом числа X-хромосом и |
сигнала от xol-1 является ген sdc-2, который вместе с |
||||||||||||
аутосом. Черви с отношением |
sdc-1 и sdc-3 контролирует ген her-1 на уровне |
||||||||||||
X:A=0.75-1.0 |
являются |
транскрипции. Ветвь генных |
взаимодействий, |
||||||||||
гермафродитами, особи, имеющие |
контролирующих |
дозовую |
компенсацию |
è |
|||||||||
баланс X:A=0.5-0.67 являются |
начинающаяся после генов sdc, на рисунке не показана |
||||||||||||
самцами. |
|
|
|
|
|
(Èç: Meyer, 1997, p. 220). |
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
346