

Синтез гема.
Исходными веществами для синтеза гема служат глицин сукцинил-КоА.
На первой стадии из
глицина и сукцинил-КоА при участии
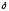 -аминолевуленатсинтазы
образуется
-аминолевуленатсинтазы
образуется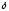 -аминолевуленовая
кислота, это процесс идет в митохондриях.
Активность фермента регулируется по
принципу обратной отрицательной связи
– гемом.
-аминолевуленовая
кислота, это процесс идет в митохондриях.
Активность фермента регулируется по
принципу обратной отрицательной связи
– гемом.
СООН СН2-NH2 HSKoA COOH
|
| ПФ |
СН2
+ COOH
CH2
+ CO2

| 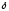 -аминолевули-
|
-аминолевули-
|
СН2 натсинтаза CH2
| |
COSKoA C=O
|
CH2-NH2
Затем из 2 молекул
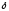 -АЛК
с участием порфобилиногенсинтазы
образуется порфибилиноген. Порфобилиноген
превращается в копропорфириногенIII,
копропорфириногенIIIпревращается через ряд стадий в
протопорфиринIX. Затем в
кольцо протопорфиринаIXвключаетсяFe2+. Затем
гем соединяется с белком глобином.
-АЛК
с участием порфобилиногенсинтазы
образуется порфибилиноген. Порфобилиноген
превращается в копропорфириногенIII,
копропорфириногенIIIпревращается через ряд стадий в
протопорфиринIX. Затем в
кольцо протопорфиринаIXвключаетсяFe2+. Затем
гем соединяется с белком глобином.
Источником Fe2+в синтезе гема является ферритин, который считается резервом гемопротеидом и откладывается в клетках костного мозга, печени и селезенки.
Установлено, что источником всех 4 атомов азота и 8 атомов углерода тетрапиррольного кольца является глицин. Источником остальных 26 из 34 атомов углерода служит сукцинил-КоА.
Талассемии – нарушение синтеза какой-либо цепи гемоглобина.
Железодефицитные анемии.
Классическим примером
наследственной гемоглобинпатии является
серповидноклеточная анемия. Глу в 6-м
положении в
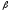 -цепи
заменен на вал. Эритроциты в условиях
низкого парциального давления кислорода
принимают форму серпа. Такой гемоглобин
после отдачи кислорода превращается в
плохо растворимую форму и начинает
выпадать в осадок в виде веретенообразных
кристаллоидов, которые деформируют
клетку и вызывают массивный гемолиз.
-цепи
заменен на вал. Эритроциты в условиях
низкого парциального давления кислорода
принимают форму серпа. Такой гемоглобин
после отдачи кислорода превращается в
плохо растворимую форму и начинает
выпадать в осадок в виде веретенообразных
кристаллоидов, которые деформируют
клетку и вызывают массивный гемолиз.
Талассемии –
генетически обусловленной нарушение
синтеза одной из нормальных цепей
гемоглобина. Угнетение синтеза
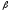 -цепей
вызывает развитие
-цепей
вызывает развитие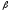 -талассемии,
угнетение синтеза
-талассемии,
угнетение синтеза -цепей
-
-цепей
- -талассемия.
При
-талассемия.
При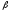 -талассемии
появляется до 15%HbA2,
повышается до 15-60% содержание фетального
гемоглобина. Болезнь характеризуется
гиперплазией и разрушением костного
мозга, поражением печени, деформацией
черепа и тяжелой гемолитической анемией.
Эритроциты имеют мишеневидную форму.
Механизм изменения формы эритроцитов
не выяснен. Название связано с тем, что
возникает у людей, живущих на побережье
Средиземного моря.
-талассемии
появляется до 15%HbA2,
повышается до 15-60% содержание фетального
гемоглобина. Болезнь характеризуется
гиперплазией и разрушением костного
мозга, поражением печени, деформацией
черепа и тяжелой гемолитической анемией.
Эритроциты имеют мишеневидную форму.
Механизм изменения формы эритроцитов
не выяснен. Название связано с тем, что
возникает у людей, живущих на побережье
Средиземного моря.
Распад гема
За сутки в организме распадается около 9 г гемопротеидов. Период жизни эритроцитов 120 дней, разрушаются они в кровеносном русле или в селезенке. Гемоглобин связывается с гаптоглобином и в виде комплекса гаптоглобин-гемоглобин поступает в клетки ретикулоэндотелиальной системы селезенки. Комплекс гаптоглобин-гемоглобин распадается и гаптоглобин переходит в кровь, а гемоглобин окисляется в метгемоглобин (Fe3+).
В РЭС селезенки гемоглобин под действием гемоксигеназы превращается в вердоглобин. Вердоглобин теряет Fe, которое связывается трансферином и доставляется кровью в костный мозг. Вердоглобин отдает белок глобин и превращается в биливердин. При восстановлении биливердина НАДФ Н2образуется билирубин.
Билирубин – плохо растворимое соединение и в крови связывается с альбумином. В виде комплекса альбумин-билирубин идет транспорт билирубина кровью в клетки печени. В печени билирубин соединяется с глюкуроновой кислотой с образованием моно (20%) и диклюкуронидов (80%), они хорошо растворимы в воде. Этот вид билирубина называется конъюгированным билирубином (связан с глюкуроновой кислотой), а также называется связанным прямым, т.к. может быть прямо обнаружен с помощью реактива Эрлиха.
Билирубинглюкурониды в незначительных количествах диффундируют в кровеносный капилляр. В плазме крови присутствуют 2 формы билирубина: неконъюгированный (непрямой, свободный) и конъюгированный (прямой, связанный) – 25% от общего билирубина. Билирубинглюкурониды с желчью поступают в кишечник, где от них отщепляется глюкуроновая кислота и вновь образуется неконъюгированный билирубин. В тонком кишечнике небольшая часть билирубина может всосаться и через портальную вену вновь поступать в печень. Остальной билирубин подвергается действию кишечных бактерий и в тонком кишечнике билирубин превращается в уробилиноген. Уробилиноген всасывается в тонком кишечнике и через воротную вену поступает в печень, где уробилиноген разрушается до моно- дипирролов.
Не разрушенный уробилиноген вновь с желчью поступает в кишечник и восстанавливается до стеркобилиногена (бесцветен). Стеркобилиноген окисляется до стеркобилина и выделяется с фекалиями. Небольшое количество стеркобилиногена поступает в почки, затем окисляется до стеркобилина и выделяется с мочой.
В норме содержание общего билирубина в сыворотке крови составляется 8-20 мкмоль/л.
Билет 95
АЗОТ ОСТАТОЧНЫЙ- азот небелковых соединений (мочевины, аминокислот, мочевой кислоты, креатина и креатинина, аммиака, индикана и др.), остающихся в сыворотке крови после осаждения белков. Концентрация А. о. в сыворотке крови является ценным диагностическим показателем при многих заболеваниях. А. о. определяют в надосадочной жидкости - после удаления осажденных белков сыворотки крови центрифугированием - с помощью азотометрического метода Кьельдаля в его многочисленных модификациях, колориметрических и гипобромитных методов. Метод Кьельдаля заключается в осаждении белков трихлоруксусной кислотой, минерализации надосадочной жидкости в присутствии концентрированной серной кислоты, перегонке образовавшегося аммиака и его количественном определении. Метод Кьельдаля в наст, время используется для проверки точности других методов определения азота. Для серийных определений он мало пригоден из-за своей трудоемкости.
В норме концентрация А. о. в сыворотке крови равна 20-40 мг/100 мл, или 14,3-28,6 ммоль/л. Увеличение содержания А. о. в крови происходит при почечной недостаточности, а также при сердечной недостаточности, злокачественных опухолях, инф. болезнях (в результате усиления распада тканевых белков и повышения содержания в крови небелковых азотсодержащих соединений). Уменьшение концентрации А. о. отмечают при беременности.
Азотемия (азот + греч. haima кровь) — избыточное содержание в крови азотсодержащих продуктов белкового обмена — см. Почечная недостаточность.
В плазме присутствуют также безазотистые органические вещества: глюкоза, нейтральные жиры и липоиды (липиды
Билет 96.
Согласно современным предтавлениям, все клеточные и внутриклеточные мембраны устроены сходным образом: основу мембраны составляет двойной молекулярный слой липидов (липидный бислой) на котором и в толще которого находятся белки ( см. рис. 1). Липидные бислои образуются амфифильными молекулами фосфолипидов и сфингомиелина в водной фазе. Амфифильными эти молекулы называют потому, что они состоят из двух частей, различных по своей растворимости в воде: полярной "головки", обладающей высоким сродством к воде, т. е. гидрофильной, и "хвоста", образуемого неполярными углеводородными цепями жирных кислот; эта часть молекулы обладает низким сродством к воде, т. е. гидрофобна.
В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин. Например, в мембранах эритроцитов человека их содержание, составляет, соответственно 36, 30 и 22 % по весу; еще 12% приходится на гликолипиды
Примером амфифильной молекулы может служить молекула фосфатидилэтаноламина, структура которой показана на рис. 2. Как и другие фосфолипиды, фосфатидилэтаноламин, в химическом отношении представляет собой сложные эфиры трехатомного спирта глицерина с двумя жирными кислотами; к третьей гидроксильной группе присоединен ортофосфат, а к нему - небольшая органическая молекула, характерная для каждого вида фосфолипидов. В рассматриваемом случае это этаноламин, но могут быть также холин, инозитол, серин и некоторые другие молекулы.
В состав липидного слоя мембран входят также холестерин и сфингомиелины; последние близки к фосфолипидам по химическому строению и физическим свойствами.
Белки мембран принято делить на интегральные и периферические. Интегральные белки имеют обширные гидрофобные участки на поверхности и нераствориммы в воде.
С липидами мембран они связаны гидрофобными взаимодействиями и частично погружены в толщу липидного бислоя, а зачастую и пронизывают бислой, оставляя на поверхности сранительно небольшие гидрофильные участки.
Отделить эти белки от мембраны удается только с помощью детергентов, типа додецилсульфата или солей желчных кислот, которые разрушают липидный слой и переводят белок в растворимую форму (солюбилизируют его) образуя с ним ассоциаты. Все дальнейшие операции по очистке интегральных белков осуществляются также в присутствии детергентов.
Периферические белки связаны с поверхностью липидного бислоя электростатическими силами и могут быть отмыты от мембраны солевыми растворами.
|
Клетки |
Мембраны |
Функции |
|
Все клетки |
Клеточные (цитоплазматические) |
Активный перенос K+, Na+, Ca2+ Подержание осмотического равновесия |
|
Большинство клеток |
Клеточные |
Связывание гормонов и включение механизмов внутриклеточной сигнализации |
|
Нервные и мышечные клетки |
Клеточные |
Генерация потенциалов покоя и действия Распространение потенциала действиЯ |
|
Большинство клеток (кроме эритроцитов) |
Внутренняя мембрана митохондрий |
Перенос электронов на кислород и синтез АТФ (окислительное фосфорилирование) |
|
Большинство клеток (кроме эритроцитов) |
Эндоплазматический ретикулум |
Перенос ионов кальция из клеточного сока внутрь везикул |
|
Клетки зрительного эпителия |
Мембраны зрительных дисков |
Поглощение квантов света и генерация внутриклеточного сигнала |
В таблице 1 перечислены функции цитоплазматических и некоторых внутриклеточных мембран. Во всех живых клетках биологические мембрану выполняют функцию барьера, отделяющего клетку от окружающей среды, и разделяющего внутренний объем клетки на сравнительно изолированные "отсеки" (compartments).
Сами по себе перегородки, разделяющие клетки на отсеки, построены из двойного слоя липидных молекул (называемого часто липидным бислоем) и практически непроницаемы для ионов и полярных молекул, растворимых в воде.
Но в этот липидный бислой встроены многочисленные белковые молекулы и молекулярные комплексы, одни из которых обладают свойствами селективных (т. е. избирательных) каналов для ионов и молекул, а другие - насосов, способных активно перекачивать ионы через мембрану. Барьерные свойства мембран и работа мембранных насосов создают неравновестное распределение ионов между клеткой и внеклеточной средой, что лежит в основе процессов внутриклеточной регуляции и передачи сигналов в форме электрического импульса между клетками.
Вторая функция, общая для всех мембран - это функция "монтажной платы" или матрицы, на которой располагаются в определенном порядке белки и белковые ансамбли, образующие системы переноса электронов, запасания энергии в форме АТФ, регуляции внутриклеточных процессов гормонами, поступающими извне и внутриклеточными медиаторами, узнавания других клеток и чужеродных белков, рецепции света и механических воздействий и т
Гибкая и эластичная пленка, которой по существу являются все мембраны, выполняет и определенную механическую функцию, сохраняя клетку целой при умеренных механических нагрузках и нарушениях осмотического равновесия между клеткой и окружающей средой.
Общие для всех мембран функции барьера для ионов и молекул и матрицы для белковых ансамблей обеспечиваются главным образом липидным бислоем, который устроен в принципе одинаково во всех мембранах.
Однако набор белков индивидуален для каждого типа мембран, что позволяет мембранам участвовать в выполнении самых разных функций в различных клетках и клеточных структурах. Некоторые из этих фукнкций упомянуты в
Самосборка фосфолипидных молекул в липидных везикулы в водном растворе. Каждая фосфолипидная молекула состоит из полярной группы и жирнокислотных хвостов. В водном растворе происходит самосборка мембран (справа) и замыкание мембран с образованием липидных пузырьков, называемых липосомами (слева).
Резюме
Биологические мембраны, наряду с цитоскелетом, формируют структуру живой клетки. Клеточная или цитоплазматическая мембрана окружает каждую клетку. Ядро окружено двумя ядерными мембранами: наружной и внутренней. Все внутриклеточные структуры: митохондрии, эндоплазматический ретикулум, аппарат Гольджи, лизосомы, пероксисомы, фагосомы, синаптосомы и т представляют собой замкнутые мембранные везикулы (пузырьки). Каждый тип мембран содержит специфический набор белков - рецепторов и ферментов; но основа любой мембраны - бимолекулярный слой липидов (липидный бислой), который во всякой мембране выполняет две главные функции: барьера для ионов и молекул и структурной основы ( матрицы) для функционирования рецепторов и ферментов.
Термин "мембраны" как окружающей клетку невидимой плёнки, служащей барьером между содержимым клетки и внешней средой и одновременно - полупроницаемой перегородкой, через которую могут проходить вода и некоторые растворенные в ней вещества, был впервые использован, по-видимому, ботаниками фон Молем и независимо К. фон Негели (1817-1891) в 1855 г для объясненеия явлений плазмолиза. В 1877 г. ботаник В. Пфеффер (1845-1920) опубликовал свой труд “Исследования осмоса” (Leipzig), где постулировал существование клеточных мембран, основываясь на сходстве между клетками и осмометрами, имеющими искусственные полупроницаемые мембраны, которые были приготовлены незадолго до этого М. Траубе.
Дальнейшее изучение осмотических явлений в растительных клетках датским ботаником Х. де Фризом (1848-1935) послужило фундаментом при создании физико-химических теорий осмотического давления и электролитической диссоциации датчанином Я. Вант-Гоффом (1852-1911) и шедским ученым С. Аррениусом (1859-1922 ). В 1888 году немецкий физико-химик В. Нернст (1864-1941) вывел уравнение диффузионного потенциала. В 1890 году немецкий физико-химик и философ В. Оствальд (1853-1932) обратил внимание на возможную роль мембран в биоэлектрических процессах.
Между 1895 и 1902 годами Э. Овертон (1865-1933) измерил проницаемость клеточной мембраны для большого числа соединений и показал прямую зависимость между способностью этих соединений проникать через мембраны и их растворимостью в липидах. Это было чётким указанием на то,что именно липиды формируют плёнку, через которую проходят в клетку вещества из окружающего раствора. В 1902 году Ю. Бернштейн (1839-1917) привлек для объяснения электрических свойств живых клеток мембранную гипотезу.
В 1925 году Гортер и Грендел показали, что площадь монослоя липидов, экстрагированных из мембран эритроцитов, в два раза больше суммарной площади эритроцитов. Гортер и Грендел экстрагировали липиды из гемолизированных эритроцитов ацетоном, затем выпаривали раствор на поверхности воды и измеряли площадь образовавшейся мономолекулярной пленки липидов. На основе результатов этих исследований было сделано предположение, что липиды в мембране располагаются в виде бимолекулярного слоя. Это предположение подтвердили исследования электрических параметров биологических мембран (Коул и Кёртис, 1935 год): высокое электрическое сопротивление, порядка 107 Омм2 и большая электроемкость 0,51 '/м2.
Вместе с тем имелись экспериментальные данные, которые свидетельствовали о том, что биологическая мембрана содержит в своем составе и белковые молекулы. Эти противоречия экспериментальных результатов были устранены Даниелли и Давсоном, предложившими в 1935 году так сказать "бутербродную" модель строения биологических мембран, которая с некоторыми несущественными изменениями продержалась в мембранологии в течении почти 40 лет. Согласно этой модели, на поверхности фосфолипидного бислоя в мембранах располагаются белки.
Билет 97.
Биохимия печени
Печень самый крупный из паренхиматозных органов. Она выполняет ряд ключевых функций.
1) Принимает и распределяет вещества, поступающие в организм из пищеварительного тракта, которые приносятся с кровью по воротной вене. Эти вещества проникают в гепатоциты, подвергаются химическим превращениям и в виде промежуточных или конечных метаболитов поступают в кровь и разносятся в другие органы и ткани.
2) Служит местом образования желчи.
3) Синтезирует вещества, которые используются в других тканях.
4) Инактивирует экзогенные и эндогенные токсические вещества, а также гормоны.
Роль печени в метаболизме углеводов
Печень играет ведущую роль в поддержании физиологической концентрации глюкозы в крови. Из общего количества поступающей из кишечника глюкозы печень извлекает ее большую часть и тратит: 10-15 % от этого количества на синтез гликогена, 60 % на окислительный распад, 30 % на синтез жирных кислот.
При физиологической гипогликемии в печени активируется распад гликогена. Первая стадия этого процесса заключается в отщеплении молекулы глюкозы и ее фосфорилировании (фермент фосфорилаза). Далее Глю-6-Ф может расходоваться по трем направлениям:
1. по пути глтколиза с образованием пировиноградной кислоты и лактата;
2. по пентозофосфатному пути;
3. расщепляться под действием фосфотазы на глюкозу и фосфор.
Преобладает последний путь, который приводит к выбросу в общий кровоток свободной глюкозы.
В печени активно протекает глюконеогенез, при котором предшественниками глюкозы являются пируват и аланин (поступающий из мышц), глицерол - из жировой ткани и с пищей ряд глюкогенных АК. Избыточное поступление глюкозы с пищей увеличивает в гепатоците интенсивность всех путей ее превращения. Так активируется ее окисление с образованием большого количества пирувата. Для его дальнейшего окисления необходимо также большое количество КоА, который также используется и для окисления жирных кислот. В результате окисление жирных кислот и распад липидов в жировых депо замедляется.
Метаболизм липидов
В печени синтезируются желчные кислоты, при дефиците которых переваривания жиров практически не происходит. В регуляции метаболизма липидов печени принадлежит ведущая роль. Так, при дефиците основного энергетического материала - глюкозы, в печени активируется окисление жирных кислот. В условиях избытка глюкозы в гепатоцитах происходит синтез триглицеридов и фосфолипидов из жирных кислот, которые поступают в печень из кишечника.
Печени принадлежит ведущая роль в регуляции обмена холестерола. Исходное вещество в его синтезе - ацетил-КоА. Т. е. Избыточное питание стимулирует образование холестерола.
В печени синтезируются транспортные формы липопротеинов.
В печени, кроме того, синтезируются кетоновые тела, в частности ацетоацетатигидрооксимаслянная кислота, которые разносятся кровью по организму.Сердечная мышца и корковый слой надпочечников предпочитают в качестве источника энергии использовать именно эти соединения, а не глюкозу.
Билет 98
Метаболизм белков
Печень использует АК, поступающие из пищеварительного тракта для синтеза собственных белков, но большая их часть идет на синтез белков плазмы крови. В печени синтезируются фибриноген, альбумины, a- иb-глобулины и липопротеиды. В печени синтезируется также т.н. лабильный резервный белок, который является как бы запасом АК, которые затем могут использоваться различными органами и тканями по мере необходимости.
Печень занимает центральное место в обмене АК, т.к. в ней активно протекают процессы их химической модификации. Кроме того, именно в печени происходит синтез мочевины.
Детоксицирующая функция печени
Детоксикация ядовитых метаболитов и чужеродных соединений (ксенобиотиков) протекает в гепатоцитах в две стадии. Реакции первой стадии катализируются монооксигеназной системой, компоненты которой встроены в мембраны эндоплазматического ретикулума. Реакции окисления, восстановления или гидролиза являются первой стадией в системе выведения из организма гидрофобных молекул. Они превращают вещества в полярные водорастворимые метаболиты.
Основной фермент гемопротеид цитохромы Р-450. К настоящему времени выявлено множество изоформ этого фермента и отнесено, в зависимости от их свойств и выполняемых функций, к нескольким семействам. У млекопитающих идентифицировано 13 подсемейств цх Р-450, условно считается, что ферменты семейства I-IV участвуют в биотрансформации ксенобиотиков, остальные метаболизируют эндогенные соединения (стероидные гормоны, простатагландины, жирные кислоты и др.).
Важным свойством цх Р-450 является способность к индукции под действием экзогенных субстратов, что легло в основу классификации изоформ в зависимости от индуцируемости тем или веществом определенной химической структуры.
На первой стадии биотрансформации происходит образование или высвобождение гидрокси-, карбоксильных, тиоловых и аминогрупп, которые являются гидрофильными, и молекула может подвергаться дальнейшему превращению и выведению из организма. В качестве кофермента используется НАДФН. Кроме цх Р-450, в первой стадии биотрансформации принимают участие цх b5и цитохромредуктаза.
Многие лекарственные вещества, попадая в организм, превращаются на первой стадии биотрансформации в активные формы и оказывают необходимый лечебный эффект. Но часто ряд ксенобиотиков не детоксицируется, а наоборот токсифицируется с участием монооксигеназной системы и становится более реакционноспособным.
Продукты метаболизма чужеродных веществ, образовавшихся на первой стадии биотрансформации, подвергаются дальнейшей детоксикации с помощью ряда реакций второй стадии. Образующиеся при этом соединения менее полярны и в связи с этим легко удаляются из клеток. Преобладающим является процесс конъюгации, катализируемый глутатион-S-трансферазой, сульфотрансферазой и UDP-глюкуронилтрансферазой. Конъюгацию с глутатионом, приводящую к образованию меркаптуровых кислот, принято рассматривать в качестве основного механизма детоксикации.
Глутатион(ведущий компонент редокс-буфера клетки) представляет собой соединение, содержащее реактивную тиоловую группу. Большая его часть находится в восстановленной форме (GSH) и играет центральную роль в инактивации токсических и реактивных продуктов. Восстановление окисленного глутатиона осуществляет фермент - глутатионредуктаза, используя как кофермент НАДФН. Коньюгаты с глутатионом, серной и глюкуроновой кислотами выводятся из организма преимущественно с мочой.
Зональность ферментных систем
Зональность метаболических комплексов печени, основного органа поддержания химического гомеостаза, определяет различие в ферментном составе между гепатоцитами перивенозной (центральной) и перипортальной (периферической) зон ацинуса (рис.4.16.1.). Это связано с их неодинаковой потребностью в кислороде различных ферментных систем.
Так, наибольшая концентрация ферментов ЦТК, катаболизма амино- и жирных кислот, цикла мочевины, глюконеогенеза отмечена в перипортальной зоне, получающей более оксигенированную кровь. Поскольку компоненты реакций второй фазы биотрансформации локализованы в клетках этой зоны ацинуса, то они более защищены от действия токсических продуктов. В гепатоцитах перицентральной зоны более активен гликолиз и первая стадия биотрансформации ксенобиотиков.
Билет 94
 1-фетоглобулин=
1-фетоглобулин= 1-фетопротеин
– белок плода. Синтезируется в печени,
концентрация 8 – 9 г/л. функция –
поддержание онкотического давления
крови. В норме здоровых людей не
содержится. Содержание увеличивается
при остром гепатите, циррозе печени,
гепатоме, беременности. Повышение
концентрации – ранний признак рака
печени.
1-фетопротеин
– белок плода. Синтезируется в печени,
концентрация 8 – 9 г/л. функция –
поддержание онкотического давления
крови. В норме здоровых людей не
содержится. Содержание увеличивается
при остром гепатите, циррозе печени,
гепатоме, беременности. Повышение
концентрации – ранний признак рака
печени.
 2
- глобулины
2
- глобулины
Гаптоглобин составляет
25% от всех
 2-глобулинов.
Гаптоглобины синтезируются в печени,
состоят из двух легких и двух тяжелых
цепей. М.М. 200 – 250 кД, концентрация 1 – 3
г/л.
2-глобулинов.
Гаптоглобины синтезируются в печени,
состоят из двух легких и двух тяжелых
цепей. М.М. 200 – 250 кД, концентрация 1 – 3
г/л.
Разновидности Нр 1-1 (азиаты)
2-1
2-2
отличаются по набору полипептидных цепей и последовательности аминокислот.
Функция – гаптоглобин образует стабильный комплекс с гемоглобином, появляющимся в плазме крови в результате внутрисосудистого гемолиза эритроцитов. Гаптоглобин – гемоглобиновый комплекс поглощается клетками РЭС, где глобин и гем подвергаются распаду, а освободившееся железо используется для синтеза гема. Такой механизм предотвращает повреждение почек гемоглобином и потерю железа с мочой. Ингибитор протеаз, транспорт В12.
Низкий уровень гаптоглобина приводит к гемолитической анемии.
Концентрация увеличивается при воспалении, инфекции, ревматической лихорадке, метастазирующем раке.
Церулоплазмин– белок голубого цвета из-за содержания меди. М.М. 150 кД, концентрация 2 – 3 г/л, синтезируется гепатоцитами. Церулоплазмин участвует в обмене меди, регулирует уровень меди в печени, участвует в обмене витамина С, адреналина, диоксифенилаланина, серотонина, мелатонина. Уменьшение содержания цероплазмина при гепатоцеребральной дистрофии (болезнь Вильсона-Коновалова) сопровождается накоплением меди в нервной ткани и печени (врожденное нарушение). Увеличение содержания церулоплазмина специфично для меланомы и шизофрении.
 2-макроглобулин– ингибитор протеаз (эндопептидаз).
М.М. 725 кД, концентрация 152 – 420 мг/100мл,
синтезируется в печени. Протеазы
появляются в крови при гибели клеток.
2-макроглобулин– ингибитор протеаз (эндопептидаз).
М.М. 725 кД, концентрация 152 – 420 мг/100мл,
синтезируется в печени. Протеазы
появляются в крови при гибели клеток.
Три белка:
 1-антитрипсин,
1-антитрипсин, 2-макроглобулин
и интер-
2-макроглобулин
и интер- -трипсиновый
ингибитор называют ингибиторами протеаз
или антипротеазами. Эти белки ингибируют
протеазы, т.е. регулируют их активность.
Антипротеазы ингибируют ферменты
свертывающей системы крови, а также
протеазы (трипсин, химотрипсин),
поступающие в кровь после гибели и
разрушения клеток.
-трипсиновый
ингибитор называют ингибиторами протеаз
или антипротеазами. Эти белки ингибируют
протеазы, т.е. регулируют их активность.
Антипротеазы ингибируют ферменты
свертывающей системы крови, а также
протеазы (трипсин, химотрипсин),
поступающие в кровь после гибели и
разрушения клеток.