

2013-СК-біохімія-09-Обмін вуглеводів-1
.pdfІ
(цит. Berg J.M. et al. Biochemistry)
І
глюкоза
АТФ
гексокіназа
АДФ
глюкоза-6-фосфат
глюкозо-6-фосфатізомераза
фруктозо-6-фосфат
АТФ
6-фосфофруктокіназа
АДФ
фруктозо-1,6-дифосфат
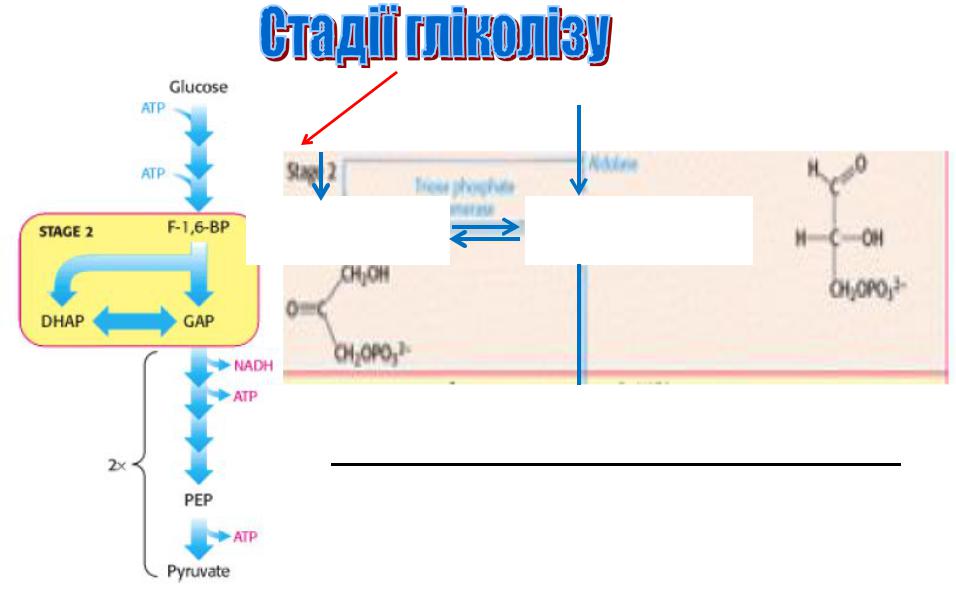
1) «Вловлювання» глюкози і її «дестабілізація»
51
(цит. Berg J.M. et al.
Biochemistry)
|
|
|
|
|
|
|
|
фруктозо-1,6-дифосфат |
|
||||
|
|
|
|
|
|
|
|
|
|
|
фруктозодифос- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
фатальдолаза |
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
ІІ |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
||
ІІ |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
тріозофосфат-ізомераза |
гліцеральдегід- |
|
|
|||||||
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
||||||||
|
|
діоксиацетон- |
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|||||||
|
|
|
фосфат |
|
|
3-фосфат |
|
|
|||||
2) |
Утворення |
двох |
взаємоперетворюваних тріоз внаслідок розщеплення фруктози (6-карбонів)
52
|
|
(цит. Berg J.M. et al. |
|
|
Biochemistry) |
|
ГА-3-Ф- |
Ф , НАД+ |
ІІІ |
наза |
н |
НАДН+Н+ |
||
|
дегідроге- |
|
|
1,3-дифосфогліцерат |
|
|
фосфоглі- |
АДФ |
|
церат- |
|
|
кіназа |
АТФ |
3-фосфогліцерат
|
|
фосфатглі |
|
|
||
|
|
|
цератму- |
|
|
|
|
2х |
|
|
таза |
|
|
|
|
|
|
|
|
|
|
|
2-фосфогліцерат |
||||
|
|
|||||
|
|
|
||||
ІІІ |
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
енолаза |
|
Н2О |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
фосфоенолпіруват
|
АДФ |
|
піруват- |
||
|
||
кіназа |
АТФ |
|
|
піруват
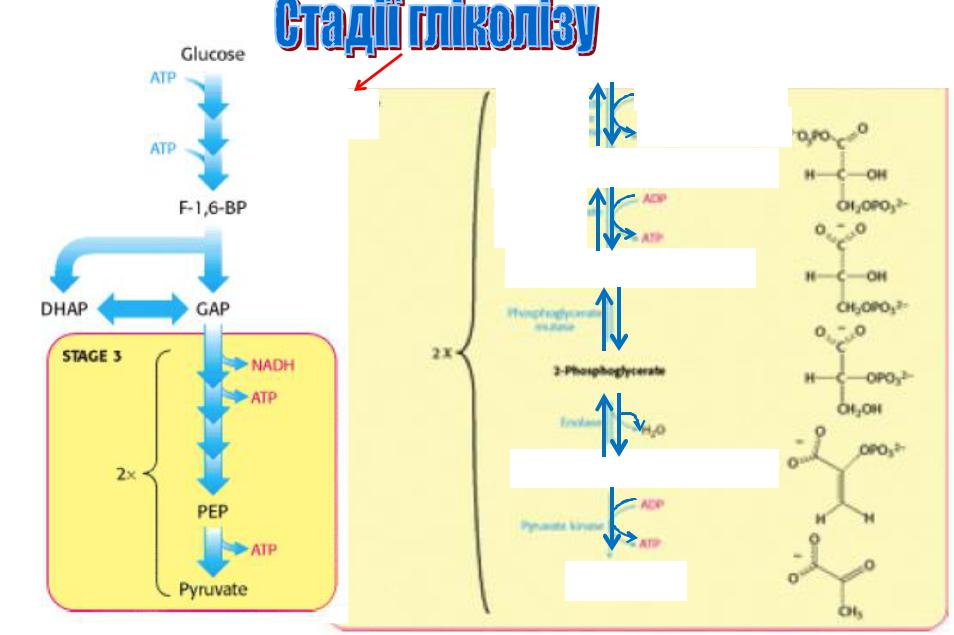
3) «Утворення» АТФ |
53 |
Сумарна реакція перетворення глюкози в піруват має такий вигляд: Глюкоза + 2Рі + 2АДФ + 2НАД+ = 2 піруват + 2АТФ + 2НАДН + 2Н+ + 2Н2О
Отже, в ході перетворення глюкози в піруват відбувається утворення двох молекул АТФ.
Гліколітичний шлях призводить до генерування АТФ в результаті розщеплення глюкози і є джерелом будівельних блоків для таких реакцій синтезу, як утворення жирних кислот з довгим ланцюгом.
Швидкість перетворення глюкози в піруват регулюється так, щоб задовольнити дві головні потреби клітини (забезпечення енергією і пластичним матеріалом)
54
Каталітичні
центри (сайти)
|
Низька [ATP] |
|
реакції |
|
|
Швидкість |
Висока [ATP] |
|
|
||
|
[фруктозо-6-фосфат] |
|
|
Фруктозо-6-фосфат |
|
|
АТФ Е |
АДФ |
Фруктозо-1, 6-дифосфат
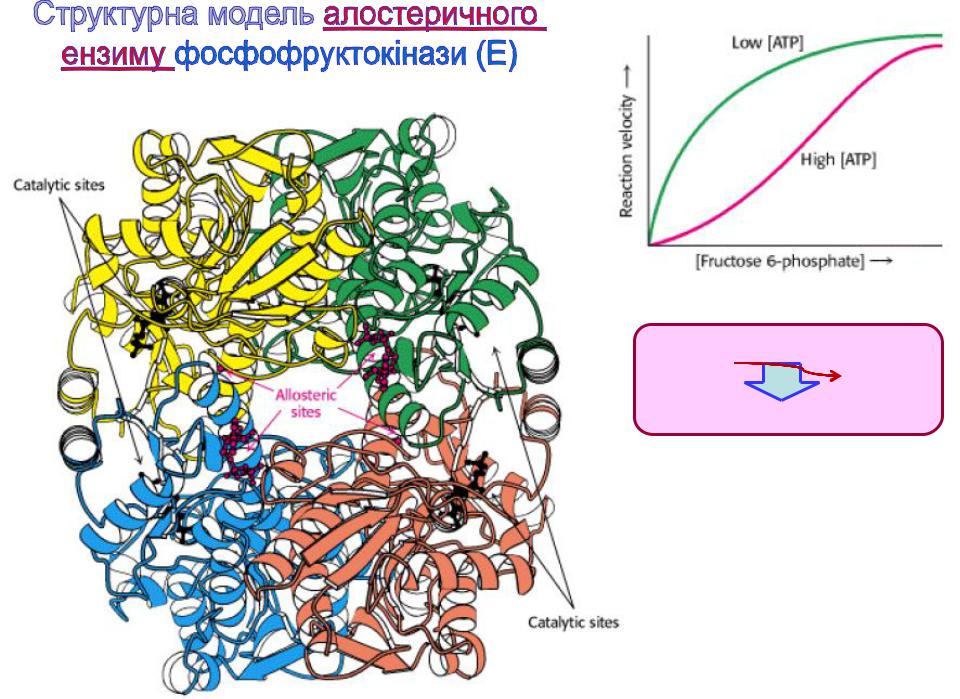
Екаталізує швидкість-
лімітуючу реакцію гліколізу.
Каталітичні
центри
(сайти)
Алостеричні центри (сайти; Allosteric sites), з якими взаємодіють алостеричні
регулятори (ефектори і
модулятори, що можуть бути як позитивними, так і негативними) 55

Контроль каталітичної активності піруваткінази
|
|
|
Pi |
|
|
|
|
|
Високий |
|
|
|
|
|
|
Низький |
|
|
фосфорильована |
глюкоза↓ |
рівень |
|||||
рівень |
глюкоза↑ |
|||||||
глюкози |
||||||||
|
неактивна |
|
|
|||||
глюкози |
|
|
|
|
у крові |
|||
|
|
піруваткіназа |
|
|
||||
у крові |
|
|
|
|
||||
|
|
|
ADP |
|
||||
|
|
|
|
|
|
|||
|
H2O |
|
|
|
|
|
||
|
|
|
|
|
|
|
||
|
Pi |
|
|
|
|
ATP |
|
|
|
дефосфорильована |
|
|
|||||
|
|
|
|
|||||
|
|
активна піруваткіназа |
|
|
||||
фосфоенолпіруват + ADP + H+ |
+ |
_ |
піруват + ATP |
|
||||
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
||
фруктозо-1,6-дифосфат ATP аланін
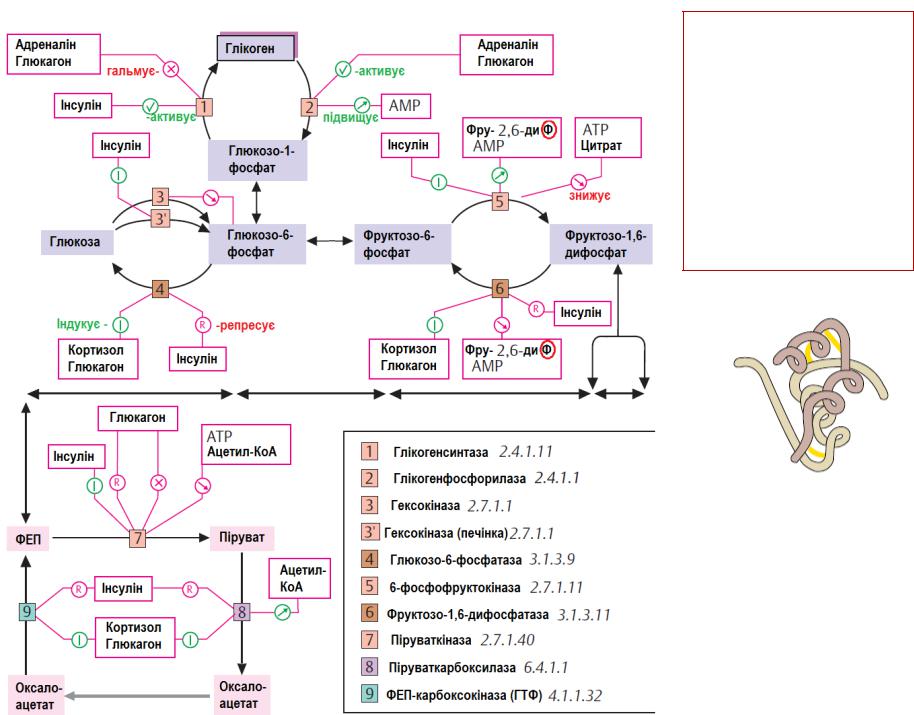
Піруваткіназа регулюється алостеричними ефекторами і ковалентною модифікацією
56
ДОАФ – диоксиацетонфосфат; 3ФГА – 3- фосфогліцериновий альдегід; 1,3-диФГ – 1,3-дифосфогліцерат; 3ФГ – 3-
фосфогліцерат; 2ФГ – 2-фосфогліцерат; ФЕП – фосфоенолпіруват.
Інсулін (51
амінокислота)
2ФГ |
3ФГ |
1,3-диФГ |
3ФГА ДОАФ |
|
Регуляція
метаболізму
вуглеводів
Адаптовано за Кольман Я.,
Рем К.-Г. Наглядная биохимия, 2009. 57

Регуляція гліколізу через вплив інгібіторів і активаторів на каталітичну активність регуляторних ензимів, які каталізують
незворотні реакції:
-гексокінази – для ферменту м'язів алостеричним інгібітором – продукт реакції глюкозо-6-фосфат;
-фосфофруктокінази – інгібіторами є метаболіт циклу Кребса цитрат і АТФ; активаторами – субстрат ензиму фруктозо-6-фосфат і АМФ.
Цей фермент каталізує швидкість-лімітуючу реакцію гліколізу.
-піруваткінази – інгібіторами є АТФ, субстрати циклу Кребса – ацетилКоА і жирні кислоти. Ізоформа піруваткінази з печінки регулюється ковалентною модифікацією - цАМФ-залежним фосфорилюванням
(дефосфорильована форма є активною, а фосфорильована –
неактивна).
У гепатоцитах піруваткіназа – індукований фермент, синтез якого стимулюється в умовах підвищеного надходження з їжею вуглеводів
та зростання рівня інсуліну.
Гальмування реакцій гліколізу за рахунок пригнічення в умовах активного клітинного дихання каталітичних активностей
фосфофруктокінази і піруваткінази є молекулярною основою ефекту
58
Пастера.
Ефект Пастера – гальмування реакцій гліколізу в умовах активації клітинного дихання
Біохімічні механізми ефекту Пастера полягають в конкуренції між цитозольною лактатдегідрогеназою та дихальним ланцюгом мітохондрій за НАДН (Н+) в окисно-відновній реакції та в зменшенні в цій метаболічній ситуації активності ключових ензимів гліколізу
– фосфофруктокінази і піруваткінази
59
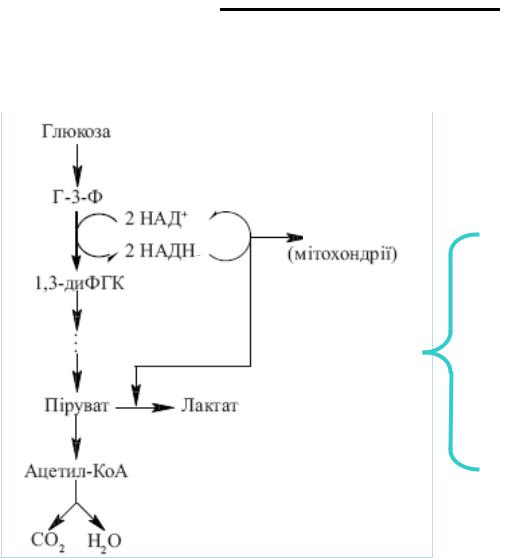
Значення анаеробного гліколізу
Анаеробний і аеробний гліколіз енергетично нерівноцінні. Утворення двох
моль лактату з глюкози супроводжується синтезом всього 2 моль АТФ, оскільки
НАДН (що утворився при окисненні гліцеральдегідфосфату) не використовується дихальним ланцюгом, а акцептується піруватом.
Не зважаючи на невеликий енергетичний ефект, анаеробний гліколіз – основне джерело енергії для скелетних м'язів у початковому періоді інтенсивної
роботи, тобто в умовах, коли постачання киснем обмежене.
|
|
|
Оскільки |
зрілі еритроцити не |
|
|
|
||
|
|
|
мають мітохондрій, то вони |
|
|
|
|
отримують |
енергію за рахунок |
|
ГА-3-Ф |
|
анаеробного окиснення. |
|
|
|
2НАДН |
|
|
|
|
|
||
Взаємовідношення між аеробним окисненням глюкози і анаеробним гліколізом
60