

Курс лекций по систематике растений_Моховидные
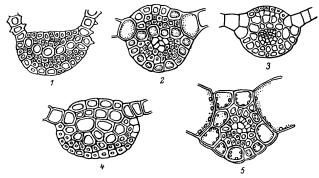
.pdfРис. 21. Форма и поперечные срезы листьев бриевых: вейссия (Weissia): 1 – лист с завёрнутыми краями; 2 - поперечный срез листа. Цинклидиум (Cinclidium): 3 – лист с сильно отвёрнутыми краями; 4 – срез через край листа. Палюделла (Paludella): 1 – оттопырено отогнутый лист. Фаброния (Fabronia): 6 – лист с реснитчатым краем; 7
– часть края листа. Гукериопсис (Hookeriopsis): 8 - широко яйцевидный лист с двойной жилкой. Эфемерум (Ephemerum) 9, 10 – листья без жилки. Индузиелла (Indusiella): 11 – лист с сильно завёрнутыми краями; 12 – поперечный срез листа. Гипнум (Hypnum): 13 – лист с двойной жилкой. Тортула (Tortula): 14 – лист с бесцветным волоском. Неккера (Neckera): 15 – языковидный поперечно-волнистый лист. Бриум (Bryum): 16 – яйцевидный лист; 17 – 18 – срезы через края листа. Гриммия (Grimmia): 19 – лист с волосковидным бесцветным кончиком.
Стебель нередко покрыт ризоидным войлоком или несет парафиллии — выросты поверхностных слоев стебля в виде зеленых пластин разной формы или сильно ветвящихся нитевидных структур. Сложноустроенных проводящих пучков в стебле нет (рис. 20). Однако нередки пучки из нескольких слабо дифференцированных клеток,
33
причем эти пучки могут формироваться или не формироваться в зависимости от условий среды.
Листья очень разнообразны по форме: яйцевидные, эллиптические, округлые, линейно-ланцетные, шиловидные, языковидные, серповидные и т.п. (рис. 21).
Листовая пластинка однослойная.
Жилка бывает простой, простой с ответвлениями, двойной, вильчатой или отсутствует. На поперечном срезе жилки различают указатели, сопроводители, спинной и брюшной стереидные пучки, спинные и брюшные клетки (рис. 22). Нередко у конкретных представителей те или иные элементы (например, брюшные клетки или сопроводители) отсутствуют.
Рис. 22. Строение жилки листа на поперечном срезе: 1 – жилка со срединными указателями, двумя стереидными пучками, спинными
ибрюшными клетками (Aulacomnium palustre); 2 – жилка со срединными указателями, спинной группой сопроводителей, двумя стереидными пучками, спинными и брюшными клетками (Mnium hornum); 3 – жилка с центральной группой сопроводителей, однородными промежуточными клетками и развитыми спинными и брюшными клетками (M. punctatum); 4 – жилка со срединными указателями, одним серповидным спинным стереидным пучком и развитыми только брюшными клетками (Tortula ruralis); 5 – жилка с двумя базальными указателями, сопроводителями, стереидным пучком
иразвитыми только спинными клетками (Funaria hygrometrica).
34
Весьма разнообразны клетки: паренхиматические (квадратные, прямоугольные, округлые, шестиугольные, ромбические, звездчатые и др.) и прозенхиматические (с различным соотношением длины и ширины); гладкие и папиллозные (или мамиллозные); окрашенные и бесцветные. Нередко клетки сильно отличаются в пределах одного листа: например, у Plagiomnium cuspidatum
большая часть листовой пластинки состоит из зеленых округлых колленхиматических клеток; край листа образован бесцветными узкими и длинными клетками, расположенными в несколько рядов и образующими кайму; на кайме имеются особые клетки-зубцы; наконец, широко и длинно низбегающие углы основания листа образованы зелеными удлиненными клетками.
Развивающиеся антеридии защищены парафизами и перигониальными листьями, которые в совокупности образуют перигоний. Архегонии также защищены парафизами и перихециальными листьями, образующими в совокупности перихеций.
После оплодотворения формируется спорогон, состоящий как обычно из трех частей: присоски, ножки и коробочки. Коробочки разнообразные по форме, обычно стегокарпные (рис. 23). Колонка полная. Перистом членистый (то есть состоит из многих клеток), простой или двойной (рис. 24). Молодой спорогон способен фотосинтезировать.
Типичные представители:
1. Верхоплодные (так называются мхи, у которых спорогон формируется на верхушке прямостоячего стебля): Dicranum polysetum (дикранум многощетинковый), Plagiomnium cuspidatum (плагиомниум остроконечный) и др.
35

Рис. 23. Различные формы коробочек и колпачков у листостебельных мхов: 1 – гомалия (Homalia); 2 – энкалипта
(Encalypta); 3 – фунария (Funaria); 4 – скоулерия (Scouleria); 5 –
сплахнум (Splachnum); 6 – ортотрихум (Orthotrichum); 7 – пелекиум
(Peleckium).
Рис. 24. Типы перистомов у бриевых мхов: 1 – тиммия
(Timmia); 2 – октоблефарум (Octoblepharum); 3 – улота (Ulota); 4 –
фонтиналис (Fontinalis); 5 – тейлория (Tayloria); 6 – цинклидиум
(Cinclidium).
36
2. Бокоплодные (так называются мхи, у которых спорогон формируется на верхушке короткой боковой веточки побега, обычно стелющегося и более или менее перисто ветвящегося): Pleurozium schreberi (плеврозиум Шребера), Hylocomium splendens (гилокомиум блестящий) и др.
Появление и основные направления эволюции моховидных
Появление моховидных тесно связано с общей проблемой происхождения высших (наземных) растений.
Наиболее вероятными предками высших растений были зеленые водоросли. Это доказывается, что у зеленых водорослей и высших растений:
1.Одинаковая ультраструктура хлоропластов.
2.Одинаковый состав фотосинтетических пигментов (хлорофилл А и В, каротиноиды).
3.Одинаковые запасные питательные вещества
(крахмал).
4.Одинаковый химический состав и структура клеточной стенки.
5.Сходное строение жгутикового аппарата у подвижных стадий.
6.Кислая рН клеточного сока.
7.Ряд других менее значимых фактов.
Для того, чтобы выдерживать хотя бы кратковременное пребывание на суше, водоросль должна иметь достаточно сложную морфологическую дифференциацию, так как одноклеточные и нитчатые водоросли переносят высыхание в виде цист и других покоящихся стадий, а следовательно не могут адаптироваться к условиям суши. По мнению F.E. Fritsch (1945) наиболее оптимальную для выхода на сушу морфоструктуру имеют водоросли порядка хетофоровых.
37
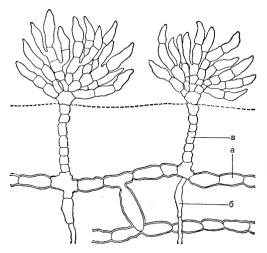
К ним, в частности, относится фричиелла клубненосная (Fritschiella tuberosa) (рис. 25), у которой имеется система подземных однорядных нитей. На них развиваются многокамерные спорангии и гаметангии, которые во влажной почве хорошо защищены от высыхания. От горизонтальных нитей отходят вертикальные, которые в воздушной среде обильно ветвятся, образуя кустики. Таким образом, у этого растения имеется как подземная часть, обеспечивающая весь организм водой и развивающая органы размножения, так и надземная, которая осуществляет фотосинтез.
Рис. 25. Строение Fritschiella tuberose: а – стелющиеся нити; б – ризоиды; в – вертикальные нити.
В настоящее время существует две группы высших растений:
1.С преобладанием в цикле воспроизводства гаметофита (моховидные);
2.С преобладанием в цикле воспроизводства спорофита (все прочие высшие растения).
38
Исходя из этого (если придерживаться монофилетической теории эволюции) предок наземных растений должен был иметь цикл воспроизводства с чередованием изоморфных поколений. Только такой цикл мог с равной вероятностью эволюционировать в противоположных направлениях. Именно такой цикл имеет фричиелла клубненосная. Это делает водоросль типа «Fritschiella tuberosa» еще более вероятным кандидатом на статус предка высших растений.
Определенную проблему представляет вопрос о местообитании гипотетического предка наземных растений. Маловероятно, чтобы это были пресноводные водоросли. Как уже упоминалось, они в силу своих биологических особенностей не способны адаптироваться к условиям суши, оставаясь в физиологически активном состоянии.
Морские водоросли, адаптированные к соленой воде, также не могут выйти на сушу, поскольку там имеются только пресноводные источники. Наиболее вероятно, что предки высших растений произрастали в устьях крупных рек (эстуариев), где дважды в сутки во время приливов-отливов соленость воды меняется от почти морской до почти пресной. Физиологические особенности таких организмов позволяют им переходить из морской воды в пресную, не теряя жизнеспособности, следовательно, именно водоросли эстуариев могли стать предками наземных растений.
Таким образом, наиболее вероятными предками высших растений были зеленые хетофоровые водоросли типа «Fritschiella tuberosa», имевшие цикл воспроизводства с чередованием изоморфных поколений и обитавшие в устьях крупных рек.
Дальнейшая вероятная история появления высших растений (моховидных и риниофитовидных) приведена на
39
Юнгерманнииды
Маршанцииды
10
9
Слоевищные растения с бесхлорофилльным и безустьичным спорогоном
8
Растения со
Политрихиды Брииды слоевищным талломом (Г =
|
|
|
|
|
|
Тетрафи |
|
|
|
|
|
|
|
|
|
|
ды |
|
|
Современ |
|
Сфагниды |
|
Андрееиды |
|
|||||||
|
|
|
|
|
|
|
|
|
ные |
7 |
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
антоцерот |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
овые |
|
|
|
|
|
|
|
|
6 |
|
||
|
Примитивные |
|
|
|||||||
|
|
|
|
|
|
|||||
|
листостебельные мхи |
|
|
|
|
|
||||
|
|
|
|
|
|
|||||
Растения с
3 Отд. Риниофиты розетковидным гаметофитом и
псилофитовидным Растения с гаметофитом спорофитом (Г ≠ С) типа такакия и
спорофитом типа
|
|
|
|
|
|
5 |
|
|
2 |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Растения с |
|
|
|
|
|
|
|
|
примитивным |
|
Растения с морфотипом |
|
|
|
|||
|
|
|
|
|
розетковидным |
||
|
такакии (Г = С) |
|
|
4 |
|
||
|
|
|
|
слоевищем из |
|||
|
|
|
|
|
|
||
|
|
|
|
|
|
|
многослойных |
|
1 |
|
|
|
|
|
пластин (Г = С) |
|
|
Растения типа |
|||||
|
|
|
|
||||
|
|
|
|
Fritchiella |
|
|
|
|
|
|
|
|
tuberosa |
|
|
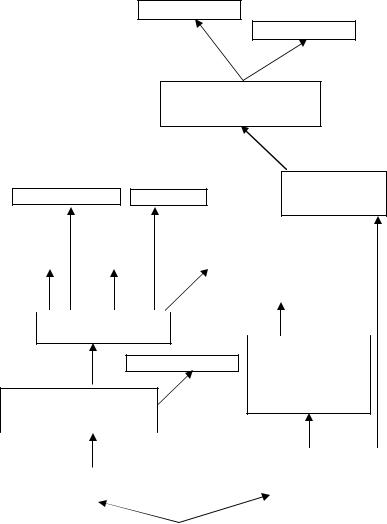
Рис. 26. Схема филогении моховидных (Г = С – гаметофит и спорофит изоморфны; Г ≠ С – гаметофит и спорофит гетероморфны).
40
рис. 26. Ниже комментируются цифры и числа, проставленные на стрелках этого рисунка.
1.Появление вальковатых осей и многоклеточных половых органов (Г = С).
2.Появление архегония и связанный с этим переход спорофита в псилофитовидное состояние с относительно быстрым отмиранием гаметофита. Появление цикла воспроизводства с чередованием гетероморфных поколений (Г ≠ С). (От этой группы растений произошли, вероятно, риниофиты).
3.Вторичная редукция спорофита и переход его на паразитическое существование на гаметофите (возможно,
врезультате неотении) (Г ≠ С). Появляется упорядоченное расположение и уплощение боковых структур, постепенно давших начало листьям. От примитивных листостебельных мхов произошли нынешние подклассы, которые в процессе эволюции приобрели характерные для них анатомо-морфологические признаки.
4.Появление многослойного розетковидного слоевища и многоклеточных половых органов (Г = С).
5.Появление архегония и связанный с этим переход спорофита в псилофитовидное состояние с быстрым отмиранием гаметофита. Появление цикла воспроизводства с чередованием гетероморфных поколений (Г ≠ С).
6.Вторичная редукция спорофита и переход его на паразитическое существование на гаметофите (возможно,
врезультате неотении) (Г ≠ С). 7. Превращение розетковидного таллома в слоевищный, благодаря редукции лишних пластин розетки.
8.Появление архегония и связанное с этим развитие зиготы на гаметофите, который не отмирал, а доминировал. При этом произошла редукция слоевища спорофита (возможно, в результате неотении) и остался
41
только спорангий, который постепенно превратился в спорогон. (Таким образом, у мхов спорогон гомологичен спорофиту, а у печеночников спорогон гомологичен спорангию).
9.Усовершенствование исходной структуры вплоть до появления слоевищных печеночников.
10.Появление листостебельного таллома из слоевищного благодаря расчленению правой и левой частей слоевища на лопасти, которые в дальнейшем обособились, стали однослойными и превратились в листья листостебельных печеночников (поэтому нет жилки). Как поверхностные выросты брюшной стороны слоевища появляются амфигастрии.
42