

35. Гипотезы о природе суммарной биоэлектрической активности мозга.
Возникновением различных форм биоэлектрической активности сопровождается любой акт жизнедеятельности – мышечное сокращение, деятельность сердца, а также деятельность мозга. Регистрация Б.Э.А. возможна с помощью спец. аппаратуры. В частности, при измерении Б.Э.А. мозга мы получаем электроэнцефалограмму мозга.
Было выдвинуто несколько гипотез о природе суммарной Б.Э.А.:
Изначально считали, что Б.Э.А. мозга создается суммой потенциалов действия клетки. Но от этой теории отказались.
Когда было выяснено, что эл. сигнал от нейрона передается через синапс, то было выдвинуто предположение, что Б.Э.А. – это сумма постсинаптических потенциалов. Это мнение и господствует в настоящее время.
Также высказывалось предположение, что Б.Э.А. – это сумма потенциалов глиальных клеток, но эта гипотеза пока не подтверждена.
36. Сверхмедленные ритмические колебания потенциалов головного мозга— вид суммарной электрической активности, представляющий собой колебания потенциала, близкие по форме к синусоидальным и имеющие частоту 6—8 в 1 мин при амплитуде 0,3—0,8 мВ (декасекундный ритм) или 0,5—2 в 1 мин с амплитудой 0,5—1,5 мВ (минутный ритм). Регистрация С. р. к. п. используется при изучении функциональных состояний мозга в норме и патологии, а также при действии фармакологических веществ.
Проводилось сравнительное исследование мощности и когерентности сверхмедленной электрической фазической активности мозга (СМФП) в диапазоне 0.05–0.5 Гц, и ЭЭГ (1.5–50 Гц) в состояниях покоя при открытых и закрытых глазах, до и после приема кофеина и бензодеазепинового транквилизатора – феназепама. Обнаружено, что прием кофеина и феназепама приводит к достаточно сходным по направленности изменениям картины ЭЭГ. Различия в эффектах препаратов проявляются в различной локализации изменений картины ЭЭГ. Предполагается, что разная локализация изменений в картине ЭЭГ отражает различия в поведенческом проявлении эффектов препаратов (стимулирующий или седативный эффект). Согласно СМФП-данным, различия в эффектах препаратов сопровождаются не только разной локализацией изменений картины СМФП, но и разной направленностью этих изменений. Это позволяет, в частности, предполагать, что при фармакопробах дифференциальная чувствительность СМФП как показателя чувствительности ЦНС может быть более высокой, чем при применении ЭЭГ, ввиду более выраженной связи поведенческой и электрографической динамики.
Билет 37. Развитие нервной системы в онтогенезе.
Онтогенез– процесс индивидуального развития особи (с момента оплодотворения до смерти организма).
Первоначальные деления зиготы приводят к формированию бластулы – зародыша (в виде полого шара), стенка которого образована одним слоем клеток. Затем в результате перемещения клеток бастулы превращается в двуслойную гаструлу. Возникающие при этом слои: эктодерма-наружный и энтодерма-внутренний, называют зародышевыми листками. В дальнейшем образуется третий – мезодерма. Нервная ткань за исключением клеток микроглии образуется из эктодермы.
Процесс, посредством которого часть клеток эктодермы превращается в специализированную нервную ткань, из которой позже развиваются спинной и головной мозг, называется пейруляцией или нейтральной индукцией. Определяющее событие для нейруляции – взаимодействие эктодермы и мезодермы. Индуцирующее действие оказывают также элементы хорды (основы будущего позвоночника).
В конце второй недели зародышевого развития участок эктодермы на дорсальной (спинной) стороне зародыша утолщается, образуя нервную пластинку. Клетки ее интенсивно размножаются и дифференцируются, приобретая узкую цилиндрическую форму. В результате интенсивного деления и неравномерного роста края нервной пластинки поднимаются, образуя нервные валики, между которыми лежит нервная бороздка.
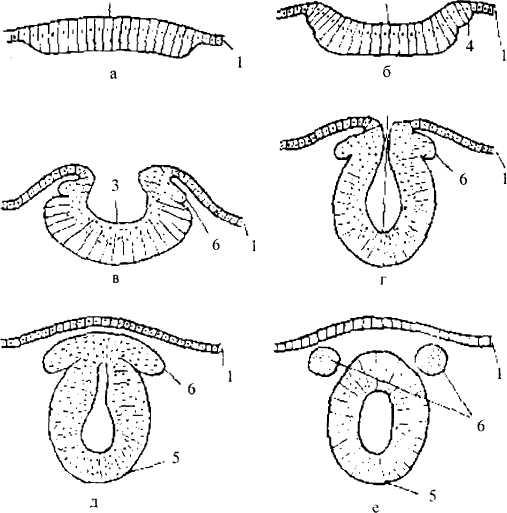
Рис. 1 /. Начальный период развития нервной системы
(поперечный разрез):
а - е — последовательные стадии развития: 1 — эктодерма:
2 — нервная пластинка; 3 — нервная бороздка; 4 — нервные вачики;
5— нервная трубка; 6— ганглиозная пластинка
Затем края нервной бороздки смыкаются, образуя нервную трубку. Она постепенно погружается в мезодерму и замыкается — сначала посередине, позже (к концу 4 недели развития) — на переднем и заднем концах. От нервной трубки по обеим ее сторонам отделяется тяж клеток — так называемая ганглиозная пластинка (нервный гребень). Ее клетки впоследствии дают элементы периферической нервной системы, паутинной и мягкой мозговых оболочек. Оставшаяся нервная трубка состоит из одного слоя клеток— нейроэпителия. В дальнейшем в результате деления этих клеток образуется слой эпендимы и слой, дающий начало нейробластам и спонгиобластам. Из нейробластов образуются нервные клетки, из спонгиобластов — глиальные клетки. Нейробласты с определенного момента прекращают делиться и начинают образовывать отростки, т.е. постепенно принимают вид зрелого нейрона. Вслед за ростом отростков следует их миелинизация и образование синапсов.
В конце 3 недели развития ростральный (головной) конец нервной трубки преобразуется в мешковидное расширение, дающее начало головному мозгу, а каудальный (хвостовой) отдел дает начало спинному мозгу. Головной конец нервной трубки скоро подразделяется на три расширения — три первичных мозговых пузыря (стадия трех мозговых пузырей). Полости этих пузырей, несколько изменяя форму, сохраняются во взрослом мозгу в виде мозговых желудочков и мозгового водопровода. Самым ростральным пузырем является первичный передний мозг, за ним следует первичный средний мозг, последний пузырь, за которым уже начинается спинной мозг, это первичный задний мозг.
На 2 месяце развития первый и третий первичные мозговые пузыри с помощью борозд разделяются, образуя каждый по два вторичных мозговых пузыря (стадия пяти мозговых пузырей). Первичный передний мозг делится на конечный мозг (полушария большого мозга и базальные ядра) и промежуточный мозг. С каждой стороны промежуточного мозга вырастает глазной пузырь, формирующий нервные элементы сетчатки глаза. Глазной бокал, образованный этим выростом, вызывает изменения в лежащей непосредственно над ним эктодерме, что приводит к возникновению из нее хрусталика. Первичный задний мозг разделяется на собственно задний мозг, включающий мозжечок и варолиев мост, и продолговатый мозг. Средний мозг сохраняется как единое целое.
Отдельные части нервной трубки растут с разной скоростью. В результате этого одновременно с формированием пяти мозговых пузырей образуются изгибы зачатка головного мозга (рис. 12). Сначала— основной изгиб в области среднего мозгового пузыря, обращенный выпуклостью дорсально; потом шейный изгиб на границе головного и спинного мозга, также выпуклостью дорсально; последний — мостовой изгиб в области заднего мозгового пузыря выпуклостью вентрально.

Рис. 12. Развитие головного мозга
А. Стадия трех мозговых пузырей:
1 — задний мозг\ 2 — средний мозг\ 3 — передний мозг.
Б. Стадия пяти мозговых пузырей:
1 — продолговатый мозг; 2 — мост и мозжечок; 3 — средний мозг; 4 — промежуточный мозг; 5— конечный мозг
После формирования мозговых пузырей в структурах ЦНС происходят сложные процессы внутренней дифференцировки и роста.
В возрасте 10-20 недель образуются все основные отделы нервной системы. К этому моменту заканчивается миграционный период ее развития, т.е все нейроны перемещаются туда, где они будут находиться во взрослом мозгу. Полушария постепенно становятся самой большой частью нервной системы, происходит выделение основных долей, начинается образование борозд и извилин. Из оболочек в ткань мозга врастают кровеносные сосуды. В спинном мозгу формируются шейное и поясничное утолщения. Окончательный вид приобретает мозжечок.
В последние месяцы эмбрионального развития в нервной системе заканчивается формирование внутренней структуры мозга (его ядер и трактов). Начинается активная миелинизация сначала спинного (в возрасте 20 недель), а затем (в возрасте 36 - 40 недель) головного мозга, которая в основном заканчивается только к 10 - 12 годам.
Мозг новорожденного весит примерно 350 г, т.е. 10% от всего веса тела. Вес мозга взрослого человека около 1250 - 1350 г. Так как деление подавляющего большинства нервных клеток прекращается еще до рождения, увеличение массы мозга происходит за счет роста тел нейронов и их отростков, миелинизации нервных волокон, деления и роста клеток нейроглии.
Таким образом, нервная система проходит следующие стадии развития:
Стадия пролиферации – процесс деления нейробластов на внутренней стороне нервной трубки (деление нервных клеток происходит только внутриутробно)
Стадия миграции (перемещение клеток в «свою часть» нервной системы)
незрелые нейроны расселяются и находят себе постоянное место.
Стадия специализации
Превращается в нервную клетку, вырастает аксон, образуются синапсы.
Конус роста – дистальная уплощенная часть аксона, которая прокладывает путь через окружающую ткань (Конус роста также образовывается при травме аксона). В конусе роста содержится фактор роста нервов (ФРН) – это белок, способствующий росту и развитию аксона.
Билет 38. «Время рефлекса» и синаптическая задержка.
Время рефлекса – это длительность реакции от момента действия раздражителя до момента самого действия (=до начала реакции).
Время рефлекса складывается из:
Длительности возбуждения сенсорных рецепторов
Проведения возбуждения по афферентным волокнам
Синаптической передачи сигнала
Проведения возбуждения по эфферентным волокнам
Самой реакции
Основное время рефлекса приходится на синаптическую передачу возбуждения -синаптическую задержку. Синаптической задержкой называется затрата времени на процессы, происходящие от момента подхода нервного импульса к пресинаптической мембране до появления в постсинаптической мембране потенциалов. При относительно высокой скорости распространения импульса по аксону, через синапс сигнал идет медленно. В большинстве центральных нейронов синаптическая задержка составляет около 0.1-0.3 мс. После этого требуется еще время на развитие возбуждающего постсинаптического потенциала (ВПСП) и потенциала действия. Весь процесс передачи нервного импульса (от потенциала действия одной клетки до потенциала действия следующей клетки) через один синапс занимает примерно 1.5 мс. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам может составить десятые доли секунды и даже целые секунды.