

Молекулярная биология клетки. Том 1
.pdf51
чужеродных молекул, избирательно выживают и размножаются. Таким образом, развитие иммунной системы индивидуального животного, подобно эволюционному процессу, использует стратегию случайных изменений с последующим отбором.
1.3.11. Нервные клетки позволяют организму быстро адаптироваться в изменяющихся условиях [17, 18]
Наличие иммунной системы - признак, свидетельствующий о высокой организации позвоночных и отличающий их от низших животных, у которых лимфоциты отсутствуют. Нервная система, напротив, имеется практически у всех многоклеточных животных. Ее роль состоит в обеспечении еще более фундаментальной потребности - потребности в быстром адаптивном ответе на внешние события.
На протяжении многих поколений эволюция совершенствует структуру организма, доводя ее до оптимальной, и приводит ее в соответствие со средой обитания. Однако в подавляющем большинстве экологических ниш наблюдаются изменения, которые происходят слишком быстро для того, чтобы могла выработаться эволюционная адаптация. В этих условиях наиболее приспособленными окажутся организмы, способные к адаптации иного рода, не требующей генетических мутаций, но тем не менее модифицирующей поведение в соответствии с изменившимися обстоятельствами. Если последовательность изменений окружающей среды полностью предсказуема, наподобие смены дня и ночи или зимы и лета, автономные изменения организма по соответствующему расписанию могут быть генетически запрограммированы. Так, фотосинтезирующая активность принадлежащего к динофлагеллятам одноклеточного организма Gonyaulax (рис. 1-39) обнаруживает 24-часовую периодичность, поддерживающуюся, даже если организм в течение недели содержать в условиях постоянного освещения. Такие биологические часы есть и у многих других организмов, но механизм их действия все еще не разгадан.
Однако большинство изменений, происходящих в окружающей среде, не предсказуемы. Например, бактерии, обитающие в кишечнике, подвергаются нерегулярным колебаниям состава и количества доступного для них питания, и любая бактерия, способная подстраивать свой метаболизм к этим изменениям, будет иметь преимущество над бактерией, не способной к такой реакции. Поэтому бактерии приобрели способность регулировать скорость синтеза своих метаболических ферментов в зависимости от концентрации питательных веществ в окружающей среде. Для сопряжения внешнего стимула с адаптивным ответом служат
Рис. 1-39. А. Ритм свечения фотосинтезирующего простейшего Gonyaulax polyedra, соотнесенный с теоретической кривой. Средний период ритма
свечения равен 22 ч 58 мин, а отклонение для индивидуальных клеток в популяции составляет в сутки ±18 мин. Б. Микрофотография
(сканирующий электронный микроскоп) Gonyaulax. (А - из D. Njus, V.D. Gooch, J. W. Hastings, Cell Biophys., 3, 223-231, 1981; Б - с любезного разрешения John Dolge.)

52
Рис. 1-40. Сравнение гормональной и нервной сигнализации. Эндокринная клетка при стимуляции высвобождает гормоны в кровяное русло, и циркулирующие гормоны вызывают ответ в любой клетке, чувствительной к ним, независимо от того, где она находится. Нервная клетка при стимуляции генерирует потенциал действия, распространяющийся вдоль аксона и быстро запускающий высвобождение нейротрансмиттера с окончания аксона; нейротрансмиттер действует только на клетки, находящиеся в непосредственной близости от окончания аксона.
специальные управляющие молекулы (такие, как циклический AMP, сАМР).
У многоклеточного организма сигнал, связывающий восприятие с действием, должен, как правило, передаваться от одних клеток к другим. Так, подстройка метаболизма часто осуществляется гормонами, которые, будучи секретированы одной группой клеток, путешествуют в тканях и вызывают ответ в других группах клеток. Но гормону требуется много времени, чтобы пройти большое расстояние, кроме того, он при этом диффундирует во все стороны. Чтобы химический сигнал передавался быстро, он должен испускаться вблизи своей мишени, тем же путем достигается точная локализация действия сигнала. Но если химический сигнал должен испускаться вблизи мишени, то как же можно использовать такой способ связи для сопряжения восприятия с реакцией на него в удаленной части организма? Проблему решают нервные клетки. Один их конец чувствителен к химическим или физическим стимулам, а другой - продуцирует химический сигнал - нейромедиатор, действующий на другие клетки (рис. 1-40). Стимуляция одного конца клетки вызывает электрическое возбуждение, которое быстро распространяется до другого конца, где вызывает высвобождение нейромедиатора. Такое сигнальное устройство позволяет многоклеточным животным быстро реагировать на изменчивый окружающий мир, а также точно координировать активность далеко отстоящих друг от друга частей тела.
1.3.12. Связи между нервными клетками определяют тип поведения
Одиночный нейрон человека не очень отличается от одиночного нейрона червя. Преимущество нервной системы человека основано на огромном количестве входящих в нее клеток и, самое главное, на способе их соединения друг с другом, который и определяет возможности нейронов в передаче, комбинировании и интерпретации сенсорных сигналов, а также в координации сложных последовательностей действий. Чтобы понять клеточные основы эволюции нервной системы, необходимо рассмотреть механизмы, с помощью которых эмбриональные нервные
53
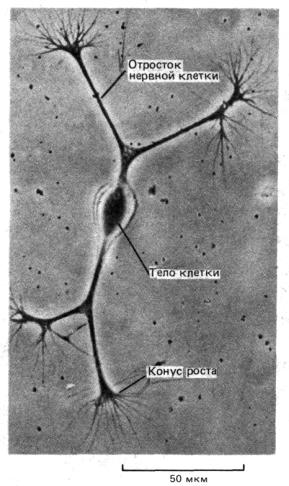
Рис. 1-41. Микрофотография изолированной нервной клетки куриного эмбриона, помещенной в культуральный сосуд с питательным раствором. У клетки появляются длинные выросты, каждый из которых продвигается с помощью структуры, называемой конусом роста. (С любезного разрешения Zoltan Gabor.)
клетки приобретают фантастически сложные формы и достигают строго упорядоченного способа их соединения (рис. 1-41). Некоторое представление об изумительной сложности и организованности сетей, которые сплетают развивающиеся нервные клетки, дает зрительная система мухи (рис. 1-42). Строение всей этой структуры задано генетически, и развитие ее происходит нормально даже в отсутствие света. При анализе развития нервной системы обнаруживается, что у всех видов оно основано на одних и тех же фундаментальных принципах: одинаковые механизмы используются для клеточного движения, межклеточной адгезии, химической сигнализации и т.д. Эволюция сложной нервной системы основывается прежде всего на эволюции сложных контрольных механизмов сочетания и координирования этих главных типов клеточного поведения.
Характер связей между нервными клетками определяет поведение животного. Без всякого обучения и опыта самец мухи спаривается с самкой, паук плетет свою паутину, птицы летят на юг. Все эти действия предначертаны в ДНК этих видов, которая контролирует поведение отдельных клеток при построении нервной системы у эмбриона и работу нервной системы у взрослого организма.
Но не всякое поведение генетически детерминировано, важное значение имеет и собственный опыт животного. Лишение развивающегося млекопитающего сенсорных стимулов может изменить микроструктуру

54
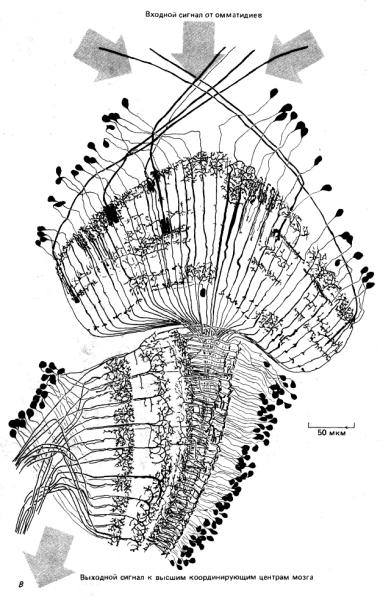
Рис. 1-42. А. Вид в сканирующем электронном микроскопе головы плодовой мушки (Drosophila). С двух сторон головы расположены два больших сложных глаза, состоящие из множества элементов, называемых омматидиями. Каждый омматидий имеет свою линзочку,
фокусирующую свет на группу находящихся в его основании клеток-фоторецепторов. Б. Схематическое изображение нейронных связей в наружном слое глаза мухи, видимых на вертикальном срезе. Свет проникает в каждый омматидий сложного глаза и фокусируется на одну из восьми находящихся в основании омматидия фоточувствительных клеток-рецепторов (здесь показаны лишь пять из них). Из-за кривизны сложного глаза свет от удаленного точечного источника фокусируется в разных омматидиях на разные фоточувствительные рецепторы. Однако короткие аксоны фоторецепторов, «смотрящих» на одну и ту же точку, переплетены таким образом, что оказываются подсоединенными к одному и тому же идущему в мозг насекомого пучку аксонов. В каждом глазу мухи присутствует более тысячи таких аксонных пучков, причем каждый из них в ходе индивидуального развития точно присоединяется к правильному набору фоторецепторов. В. Сеть нервных клеток участка мозга мухи, получающего и обрабатывающего входной сигнал от омматидиев. (А - с любезного разрешения Rudi Turner and Antony Mahowald; В - из N. Stausfeld, Atlas of an Insect Brain. New York, Springer, 1976.)
мозга. Взрослые животные почти всех видов - от кишечнополостных до человека - в той или иной степени способны к обучению. По определению обучение - это результат опыта и, следовательно, электрической активности нервных клеток, которая должна приводить к длительным изменениям нейронных связей. Детальная расшифровка этих механизмов является одной из центральных задач современных нейробиологических исследований.
Многие нейронные связи мозга, позволяющие нам читать, писать и говорить, появляются в результате обучения и отражают негенетический тип наследования. Обучение и обмен информацией дали возмож-
55
ность человеку как виду адаптироваться таким способом, который для менее высокоорганизованных существ возможен только лишь путем генетической эволюции. Тем не менее ясно, что даже эти очень сложные способности, лежащие в основе нашей культуры и общества, имеют в своей основе тонкие особенности поведения клеток - правила, по которым нейроны на долгое время модифицируют свои связи в результате электрической активности.
Конечно, изучая лишь единичные клетки, мы не поймем, как устроен многоклеточный организм, точно так же, как мы не поймем устройства клетки, анализируя отдельные биологические молекулы. Но все же, не

56
Рис. 1-43. Эволюционное родство между некоторыми упомянутыми в этой книге организмами. Ветви древа показывают пути общего происхождения, но их длина не отражает реального временного масштаба. (Отметим также, что вертикальная ось диаграммы демонстрирует не
время, а основные категории организмов.)
зная строения клетки, мы не можем рассчитывать на полное понимание работы организма. И мы не можем как следует разобраться в механизмах функционирования клетки, не зная составляющих ее молекул. Таким образом, обсуждение живой клетки, к которому мы переходим в следующей главе, должно начинаться с описания входящих в ее состав молекул.
Заключение
Эволюция крупных многоклеточных организмов связана со способностью эукариотических клеток по-разному экспрессировать наследственную информацию, а также с умением этих клеток функционировать сообща. Одним из наиболее ранних этапов на пути к многоклеточности было
57
появление эпителия, в котором клетки соединены в слои, отделяющие внутреннюю среду организма от внешнего окружения. Первыми примитивными типами дифференцированных клеток должны были быть наряду с эпителиальными клетками нервные клетки, мышечные клетки и клетки соединительной ткани. Все эти типы клеток можно найти даже у очень примитивных современных животных.
Эволюция высших животных, используя ту же основную стратегию развития, шла в направлении создания все возрастающего числа специализированных клеточных типов и все более утонченных методов координации их активности (рис. 1-43). Две системы клеток высших животных представляют каждая в своем роде вершину сложности многоклеточной организации. Одна - это иммунная система позвоночных, клетки которой способны производить миллионы различных антител. Другая - это нервная система. У низших животных большая часть нейронных связей жестко генетически детерминирована, и программа поведения эволюционирует лишь благодаря мутациям генетического материала. У высших животных работа и структура нервной системы становились все более подверженными модификациям (обучению) благодаря способности нервных клеток изменять свои связи в ответ на вызванную внешними стимулами электрическую активность.
Литература
Общая
Bendall D. S. ed. Evolution from Molecules to Men. Cambridge, UK. Cambridge University Press, 1983.
Curtis H. Biology, 4th ed. New York. Worth, 1983.
Darnell J. E., Lodish H. F., Baltimore D. Molecular Cell Biology, Chapter 25.
New York. W.H. Freeman, 1986. Darwin C. On the Origin of Species. London. Murray, 1859. Reprinted, New York, Penguin, 1984. Evolution. Sci. Am. 239(3), 1978. (An entire issue devoted to the topic.)
Keeton W. Т., Gould J. W., Gould C. G. Biological Science, 4th ed. New York, Norton, 1986. Maynard Smith J. The Theory of Evolution, 3rd ed. New York, Penguin, 1975.
Watson J. D., Hopkins N. H., Roberts J. W., Steitz J. A., Weiner A. M. Molecular Biology of the Gene, 4th ed. Chapter 28. Menlo Park CA. Benjamin-Cummings, 1987.
Wilson E. B. The Cell in Development and Heredity, 3rd ed. New York: Macmillan, 1925. (Reprinted, New York: Garland, 1987.)
Цитируемая
1.Ferris J.P., Usher D.A. Origins of life. In: Biochemistry, 2nd ed. (G. Zubay ed.), pp. 1120-1151. New York: Macmillan, 1988. Miller S. M., Orgel L. E. The Origins of Life on the Earth. Englewood Clifis, NJ. Prentice-Hall. 1974.
Schopf J. W., Hayes J. M., Walter M. R. Evolution of earth's earliest ecosystems: recent progress and unsolved problems. In: Earth's Earliest Biosphere: Its Origin and Evolution (J. W. Schopf ed.), pp. 361-384. Princeton, NJ. Princeton University Press, 1983.
2.Eigen M., Gardiner W., Schuster P., Winkler-Oswatitsch R. The origin of genetic information. Sci. Am., 244(4), 88-118, 1981.
3.Cech T.R. RNA as an enzyme. Sci. Am. 255(5), 64-75, 1986.
4.Alberts В. М. The function of the hereditary materials: biological catalyses reflect the cell's evolutionary history. Am. Zool. 26, 781 - 796, 1986. Darnell J. E., Doolittle W. F. Speculations on the early course of evolution. Proc. Natl. Acad. Sci. USA, 83, 1271 - 1275, 1986.
Orgel L.E. RNA catalysis and the origin of life. J. Theor. Biol. 123, 127-149, 1986.
5.Rogers M. et al. Construction of the mycoplasma evolutionary tree from 5S rRNA sequence data. Proc. Natl. Acad. Sci. USA, 82, 1160-1164, 1985.
6.Vidal G. The oldest eukaryotic cells. Sci. Am., 250(2), 48-57, 1984.
7.Clarke P. H. Enzymes in bacterial populations. In: Biochemical Evolution (H. Gutfreund ed.), pp. 116-149. Cambridge UK. Cambridge University Press, 1981.
58
Doolittle W.F. Archaebacteria coming of age. Trends Genet., 1, 268-269, 1985.
Wоese C. R. Bacterial evolution. Microbiol. Rev., 5, 221-271, 1987.
8.Wilson A. The molecular basis of evolution. Sci. Am., 253(4), 164-173, 1985.
9.Dickerson R. E. Cytochrome с and the evolution of energy metabolism. Sci. Am., 242(3), 136-153, 1980.
10.Cavalier-Smith T. The origin of eukaryotic and archaebacterial cells. Ann.N.Y. Acad. Sci. 503, 17-54, 1987. Margulis L. Symbiosis in Cell Evolution. New York, W. H. Freeman, 1981.
11.Vossbrinck C.R., Maddox J.V., Friedman S., Debrunner-Vossbrinck B. A., Woese C. R., Ribosomal RNA sequence suggest microsporidia are extremely ancient eukarvotes. Nature, 326, 411-414, 1987.
Yang D., Oyaizu Y., Oyaizu H., Olsen G. J., Woese C. R. Mitochondrial origins. Proc. Natl. Acad. Sci. USA, 82, 4443-4447. 1985.
12.Sleigh M. A. The Biology of Protozoa. London, Edward Arnold, 1973.
Sogin M. I., Elwood H. J., Gunderson J. H. Evolutionary diversity of eukaryotic small-subunit rRNA genes. Proc. Natl. Acad. Sci. USA, 83, 1383-1387, 1986.
13.Buchsbaum R. Animals Without Backbones, 2nd ed. Chicago. University of Chicago Press, 1976. Field K. G. et al. Molecular phytogeny of the animal kingdom. Science, 239, 748-753, 1988.
Margulis L., Schwartz K. V. Five Kingdoms: An illustrated Guide to the Phyla of Life on Earth, 2nd ed. New York. W.H. Freeman, 1987. Shapiro J. A. Bacteria as multicellular organisms. Sci. Am., 258(6), 82-89, 1988.
Valentine J. W. The evolution of multicellular plants and animals. Sci. Am., 239(3), 140-158, 1978.
14.Bode P. M., Bode H. R. Patterning in Hydra. In: Pattern Formation (G. M. Malacinski, S.V. Bryant eds.), pp. 213-244. New York, Macmillan, 1984.
15.Raff R. A., Kaufman T. C. Embryos, Genes, and Evolution. New York. Macmillan, 1983.
16.Maynard Smith J. The Evolution of Sex. Cambridge, UK. Cambridge University Press, 1978.
Michod R. E., Levin B. R. eds. The Evolution of Sex: An Examination of Current Ideas. Sunderland, MA. Sinauer, 1988.
17.Winfree A. T. The Timing of Biological Clocks. New York. W. H. Freeman, 1987.
18.Bullock Т. Н., Orkand R., Grinnell A. Introduction to Nervous Systems. San Francisco. Freeman, 1977.
59
2. Малые молекулы, энергия и биосинтез
«Хочу сообщить, что я научился получать мочевину, не используя для этого изолированную почку или животное, будь то человек или собака». Эти слова, написанные 150 лет назад молодым немецким химиком Вёлером (Wohler), ознаменовали собой конец веры в особую жизненную силу (vital force), присущую живым организмам и обусловливающую характер их отличительных особенностей. Однако то, что было откровением во времена Вёлера, сегодня звучит вполне обыденно - живые существа состоят из химических соединений. В современном представлении о жизни нет места ни для витализма, ни для чего бы то ни было, выходящего за рамки законов химии и физики. Из этого отнюдь не следует, что в биологии не осталось тайн: как покажут следующие главы, в ней еще много «белых пятен». Тем не менее следует сразу же подчеркнуть, что объем накопленных знаний поистине колоссален.
Сейчас мы располагаем подробными сведениями о важнейших молекулах клетки, причем не о каком-то небольшом их числе, а почти обо всех. Во многих случаях нам точно известна их химическая структура, так же как и пути их образования и распада. В общих чертах мы представляем себе, как химическая энергия поддерживает процессы биосинтеза в клетке, как на основе законов термодинамики создается молекулярная упорядоченность и как регулируются и координируются мириады непрерывно протекающих внутри клетки химических превращений.
В этой и следующей главах мы познакомимся вкратце с химическими свойствами живой клетки. Здесь будут рассматриваться процессы, протекающие с участием малых молекул: механизмы, с помощью которых клетка синтезирует свои главные химические компоненты и получает необходимую энергию. В гл. 3 речь пойдет о гигантских молекулах (полимерах), чьи свойства определяют специфичность биологических процессов и передачу биологической информации.
2.1. Химические компоненты клетки
2.1.1. Основа клеточной химии - соединения углерода [1]
Живая клетка состоит из ограниченного набора элементов, причем на долю шести из них (С, Н, N, О, Р, S) приходится более 99% ее общей массы. Такой состав, заметно отличающийся от состава земной коры, свидетельствует о химизме особого типа (рис. 2-1). В чем же своеобразие химии живого и как оно возникло в процессе эволюции?
Соединение, которое живая клетка содержит в наибольшем количестве, - это вода. Она составляет около 70% массы клетки, и большинство внутриклеточных реакций протекает в водной среде. Жизнь на нашей планете возникла в океане, и условия этой первобытной среды наложили неизгладимый отпечаток на химию живых существ. «Конструкция» всех живых организмов связана с уникальными свойствами

60
Рис. 2-1. Относительное количество химических элементов, обнаруженных в земной коре (неживой мир), по сравнению с количеством тех же элементов в мягких тканях живых организмов. Относительное количество выражено в процентах к общему числу имеющихся атомов. Так, например, на долю водорода приходится около 50% от числа всех атомов, присутствующих в живых организмах.
воды, такими, как полярный характер ее молекул, способность к образованию полярных связей и большое поверхностное натяжение (схема 2-1). Если не считать воды, можно сказать, что почти все молекулы клетки, за небольшим исключением, относятся к соединениям углерода,
которые рассматриваются в курсе органической химии. Среди всех элементов Земли углерод занимает особое место по способности к образованию больших молекул; до некоторой степени аналогичной способностью обладает кремний, однако он сильно уступает углероду в этом отношении. Благодаря малому размеру и наличию на внешней оболочке четырех электронов атом углерода может образовать четыре прочные ковалентные связи с другими атомами. Наиболее важное значение имеет способность атомов углерода соединяться друг с другом. образуя цепи и кольца и создавая в результате большие и сложные молекулы, на размеры которых не накладывается никаких видимых ограничений. Другие атомы, широко представленные в клетке (Н, N и О), имеют, как и углерод, небольшие размеры и способны образовать очень прочные ковалентные связи
(схема 2-2).
В принципе простые правила образования ковалентной связи между углеродом и другими элементами допускают существование астрономически большого числа соединений. Количество различных углеродных соединений в клетке действительно очень велико, но это лишь крошечная часть теоретически возможного. В некоторых случаях мы можем довольно убедительно обосновать, почему то или иное соединение выполняет именно данную биологическую функцию; однако чаще возникает ощущение, что выбор пал на один из многих приемлемых вариантов и свою роль здесь сыграл случай (рис. 2-2). Определенные типы реакций и химические мотивы, однажды установившись, сохранили (с некоторыми вариациями) свой характер в ходе эволюции. Появление новых классов соединений было, очевидно, необходимым или целесообразным лишь в редких случаях.