

Вопрос20. Структурно-функциональная характеристика глиальных клеток
Глия — структура нервной системы, образованная специализированными клетками различной формы, которые заполняют пространства между нейронами или капиллярами, составляя 10% объема мозга.
Виды глии
Различают следующие виды глии: астроглия, олигодендроглия, микроглия
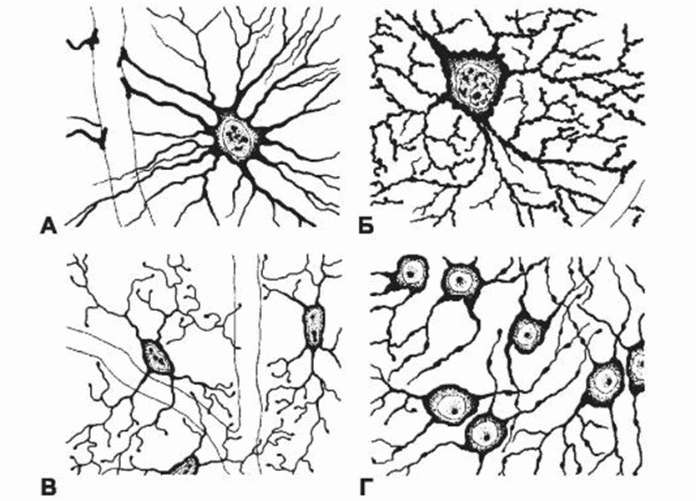
А — волокнистый астроцит; Б — протоплазматический астроцит; В — микроглия; Г — олигодендроглиоциты
Особенности глиальных клеток
Одной из особенностей глиальных клеток является их способность к изменению своего размера. Изменение размера глиальных клеток носит ритмический характер: фазы сокращения — 90 с, расслабления — 240 с, т.е. это очень медленный процесс. Средняя частота ритмических изменений варьирует от 2 до 20 в час. При этом отростки клетки набухают, но не укорачиваются в длине.
Глиальная активность изменяется под влиянием различных биологически активных веществ: серотонин вызывает уменьшение указанной «пульсации» олигодендроглиальных клеток, норадреналин — усиление. Хлорпромазин действует так же, как и норадреналин. Физиологическая роль «пульсации» глиальных клеток состоит в проталкивании аксоплазмы нейрона и влиянии на ток жидкости в межклеточном пространстве.
Физиологические процессы в нервной системе во многом зависят от миелинизации волокон нервных клеток. В центральной нервной системе миелинизация обеспечивается олигодендроглией, а в периферической — шванновскими клетками.
Глиальные клетки не обладают импульсной активностью, подобно нервным, однако мембрана глиальных клеток имеет заряд, формирующий мембранный потенциал. Его изменения медленны, зависят от активности нервной системы, обусловлены не синаптическими влияниями, а изменениями химического состава межклеточной среды. Мембранный потенциал глии равен примерно 70-90 мВ.
Глиальные клетки способны к распространению изменений потенциала между собой. Это распространение идет с декрементом (с затуханием). При расстоянии между раздражающим и регистрирующим электродами 50 мкм распространение возбуждения достигает точки регистрации за 30-60 мс. Распространению возбуждения между глиальными клетками способствуют специальные щелевые контакты их мембран. Эти контакты имеют пониженное сопротивление и создают условия для электротонического распространения тока от одной глиальной клетки к другой.
Так как глия находится в тесном контакте с нейронами, то процессы возбуждения нервных элементов сказываются на электрических явлениях в глиальных элементах. Это влияние связывают с тем, что мембранный потенциал глии зависит от концентрации К+ в окружающей среде. Во время возбуждения нейрона и реполяризации его мембраны вход ионов К+ усиливается. Это значительно изменяет его концентрацию вокруг глии и приводит к деполяризации ее клеточных мембран.
Глиальные клетки служат опорным и защитным аппаратом для нейронов. Метаболизм глиальных клеток тесно связан с метаболизмом нейронов, которые они окружают. Возможно, глиальные клетки участвуют в процессах памяти. Сателлиты нейроглии, называемые шванновскимя клетками, синтезируют оболочки миелинизированных нервных волокон периферических нервов. Некоторые глиальные клетки выполняют функции фагоцитов.
Клетки нейроглии делят на ряд типов. Эпендимоциты макроглии выстилают желудочки головного мозга и спинномозговой канал и образуют эпителиальный слой в сосудистом сплетении. Они соединяют желудочки с подлежащими тканями и выполняют разграничительную, опорную и секреторную функции. Клетки макроглии делятся на две категории -астроциты и олигодендроциты. Астроциты выполняют опорную функцию, осуществляют транспорт питательных веществ к нейронам, поглощение погибших клеток, участвуют в регулировании состава жидкости экстрацеллюлярного пространства (глюкоза, аминокислоты, ионы, в частности, являются буфером и депо ионов калия). Протоплазматические астроциты локализованы в сером веществе мозга. От тела астроцита, содержащего овальное ядро и большое количество гликогена, отходят сильно разветвленные короткие и толстые отростки. Фибриллярные астроциты локализованы в белом веществе мозга. Ядро у них также овальное, и тело клетки также содержит много гликогена, но отростки длинные и менее разветвленные. Некоторые ветви фибриллярных астроцитов буквально упираются в стенки кровеносных сосудов. Полагают, что эти клетки переносят питательные вещества из крови в нейроны. Астроциты этих двух типов взаимодействуют в обширной трехмерной сети, в которой расположены нейроны. Здесь астроциты часто делятся и в случае повреждений мозга образуют рубцовую ткань.
Олигодендроциты локализованы в сером и белом веществе мозга. Их главной функцией является изоляция миелином нейронов центральной нервной системы. Олигодендроциты мельче астроцитов и имеют одно сферическое ядро. От тела клетки отходит небольшое число тонких веточек. Цитоплазма олигодендроцитов содержит большое количество рибосом. Шванновские клетки-это специализированные олигодендроциты, синтезирующие миелиновую оболочку миелинизированных волокон.
Клетки микроглии локализованы и в сером, и в белом веществе, но в сером веществе их больше. Эти клетки продолговатой формы и содержат лизосомы и хорошо развитый аппарат Гольджи. От каждого конца тела клетки отходит толстый отросток. Отростки ветвятся мелкими боковыми веточками. При повреждениях мозга клетки микроглии превращаются в фагоциты, способные перемещаться посредством амебоидного движения.
Основные отличия глиальных клеток от нейронов:
1) Глиальные клетки имеют только один тип отростков, в то время как нейроны имеют два типа отростков - аксоны и дендриты.
2) Глиальные клетки, хотя и имеют потенциал покоя (~89 мв), не могут генерировать потенциал действия как нейроны.
3) Глиальные клетки не имеют химических синапсов как нейроны.
4) Глиальные клетки, в отличие от зрелых нейронов, способны к делению.
5) Число глиальных клеток в центральной нервной системе в 10-50 раз больше, чем нейронов.
21. Мембранный потенциал покоя и механизм его формирования
Потенциа́л поко́я — мембранный потенциал возбудимой клетки (нейрона, кардиомиоцита) в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 мВ[1]. У нейронов и нервных волокон обычно составляет -70 мВ.
Возникает вследствие диффузии отрицательно заряженных ионов хлора из окружающей среды в цитоплазму клетки в процессе установления осмотического равновесия. Анионы органических кислот, нейтрализующие заряд ионов калия в цитоплазме, не могут выйти из клетки, однако ионы хлора, концентрация которых в цитоплазме мала по сравнению с окружающей средой, диффундируют в цитоплазму до тех пор, пока создаваемый ими электрический заряд не начнёт уравновешивать их градиент концентрации на клеточной мембране.
Для того, чтобы на мембране поддерживалась разность потенциалов, необходимо, чтобы была определенная разность концентрации различных ионов внутри и снаружи клетки.
Концентрации ионов в клетке скелетной мышцы и во внеклеточной среде
Ионы Концентрация в саркоплазме (ммоль) Концентрация вне клетки (ммоль)
Ионы Концентрация в саркоплазме (ммоль)
K+ 140 2,5Концентрация вне клетки (ммоль)
Na+ 10 120
Cl- 3-4 120
Ca2+ <0,001 2
A- (полипептиды) 140 0
С помощью уравнения Нернста можно рассчитать равновесный трансмембранный потенциал для K+, который и определяет значение ПП. Но значение потенциала покоя полностью не совпадает с EK+, так как в создании его участвуют также ионы натрия и хлора, вернее, их равновесные потенциалы.
Впоследствии было доказано, что основной вклад в создание потенциала покоя вносит выходящий калиевый ток, который осуществляется через специфические белки-каналы — калиевые каналы постоянного тока. В покое калиевые каналы открыты, а натриевые каналы закрыты. Ионы калия выходят из клетки по градиенту концентрации, что создает на наружной стороне мембраны избыток положительных зарядов; при этом на внутренней стороне мембраны остаются отрицательные заряды. Некоторый (небольшой) вклад в создание потенциала покоя вносит также работа так называемого "натрий-калиевого насоса", который образован особым мембранным ферментом - натрий-калиевой АТФазой.
Потенциал покоя для большинства нейронов составляет величину порядка −60 мВ — −70 мВ. У клеток невозбудимых тканей на мембране также имеется разность потенциалов, разная для клеток разных тканей и организмов.
Формирование потенциала покоя[править | править вики-текст]
ПП формируется в два этапа.
Первый этап: создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na+ на K+ в соотношении 3 : 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ, обеспечивает его электрогенность.
Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы:
1. Дефицит ионов натрия (Na+) в клетке.
2. Избыток ионов калия (K+) в клетке.
3. Появление на мембране слабого электрического потенциала (-10 мВ).
Второй этап: создание значительной (-60 мВ) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K+. Ионы калия K+ покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до −70 мВ.
Потенциал покоя.
Когда клетка или волокно находится в состоянии покоя, ее внутренний потенциал (мембранный потенциал) варьирует от -50 до -90 милливольт и условно принимается за ноль. Наличие этого потенциала обусловлено неравенством концентраций ионов Na+,K+,Cl-,Ca2+ внутри и вне клетки, а также различной проницаемостью мембран для этих ионов. Внутри клетки калия в 30-50 раз больше, чем снаружи. При этом проницаемость мембраны невозбужденной клетки для ионов калия в 25 раз выше, чем для ионов натрия. Поэтому калий выходит из клетки наружу. В этаже время анионы цитоплазмы клетки особенно наружные хуже проходят через мембрану, концентрируются у ее поверхности, создавая «―» потенциал. Вышедшие из клетки ионы калия удерживаются у наружной поверхности мембраны электростатическим противоположным зарядом.
Это разность потенциала называется мембранным потенциалом или потенциалом покоя. Со временем при такой ситуации большинство ионов калия могли бы выйти за пределы клетки и разность концентраций их снаружи и внутри выровнялась бы, но этого не происходит, т. к. в клетке сущ-ет натрий калиевый насос. Благодаря которому осуществляется обратное поступление калия из тканевой жидкости в клетку и выделение ионов натрия против градиента концентрации (а натрия больше снаружи клетки
Итак, мембранный потенциал покоя — это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.