

2 курс / Гистология / Общий курс гистологии
.pdf6 класс – мегакариоцит – гигантская клетка, достигающая 50–80 мкм, имеет неправильной лопастевидной формы ядро и слабобазофильную цитоплазму со специфическими гранулами, содержащими факторы свертывания. Среди мегакариоцитов выделяют резервные (не образуют тромбоцитов) и зрелые (дают кровяные пластинки).
Микровезикулы цито- |
|
|
|
|
|
|
|
|||
плазмы зрелого мега- |
|
|
|
|
|
|
|
|||
кариоцита формируют |
|
|
|
|
|
|
|
|||
пограничные |
мем- |
|
|
|
|
1 |
|
|
||
браны, которые делят |
|
|
|
|
|
|
|
|||
|
21 |
|
|
|
|
|
||||
клетку на участки, со- |
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
||||
держащие до 3-х гра- |
|
|
|
|
|
|
|
|||
нул. В силу своих |
|
|
|
|
|
|
|
|||
размеров |
мегакарио- |
|
|
|
|
|
|
|
||
цит не может проник- |
|
|
|
|
|
|
|
|||
|
|
|
3 |
|
|
|||||
нуть в кровоток и по- |
|
|
|
|
|
|||||
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
||||
этому отщепляет |
эти |
|
|
|
|
|
|
|
||
участки |
цитоплазмы |
|
|
|
|
|
|
|
||
по пограничным (де- |
|
|
|
|
|
|
|
|||
маркационным) |
ли- |
|
|
|
|
|
|
|
||
ниям. Фрагменты ци- |
Рис. 28. Схема отшнуровки тромбоцитов от мега- |
|||||||||
топлазмы |
проходят |
|||||||||
кариоцита (по О.В. Волковой). 1 – гранулы и органел- |
||||||||||
через поры синусоид- |
||||||||||
лы, 2 – демаркационные линии, 3 – ядро мегакариоцита. |
||||||||||
ных капилляров. |
Так |
|
|
|
|
|
|
|
||
происходит образование кровяных пласти-
нок – тромбоцитов (рис. 28). Вокруг ядра мегакариоцита остается узкий ободок цитоплазмы. Такой мегакариоцит называется резидуальным, и он вскоре разрушается.
Иммунитет (лат. «immunistas» – в Древнем Риме гражданин, свободный от государственных повинностей) – способ защиты ор-
ганизма от всех антигенно чужеродных веществ, обеспечивающий генетическую целостность особей вида в течение их индивидуальной жизни.
Антигены – структурно чужеродные для данного конкретного организма вещества, способные вызывать иммунный ответ. Для них характерны два признака: антигенная специфичность – свойство, отличающее данный антиген от индивидуального, антигенного состава организма, в который он проник, и иммуногенность – способность стимулировать иммунную систему к формированию иммунно-
71
го ответа. Свойствами антигенов обладают бактерии, вирусы, паразиты, опухолевые клетки, чужеродные ткани, а также продукты их жизнедеятельности (например, белки, полисахариды, полипептиды).
Вещества, реагирующие с антителами, но неспособные при введении индуцировать специфический иммунный ответ, называются гаптенами. Участок молекулы антигена, способный связываться с активным центром антитела или антигенсвязывающего рецептора лимфоцита, называется антигенной детерминантой.
Антигены главного комплекса гистосовместимости – глико-
протеины плазматической мембраны, которые находятся на поверхности всех ядерных клеток организма. Свое название они получили в связи со способностью вызывать сильную реакцию отторжения при пересадке тканей в пределах одного вида. У представителей разных видов система антигенов главного комплекса гистосовместимости,
МНС (major hystocompatibility complex), имеет разные названия. У
человека она называется HLA (human leucocite antigens). В антигенном распознавании участвуют антигены МНС класса I и класса II. Антигены класса I представлены на поверхности практически всех клеток. Антигены класса II экспрессированы преимущественно на мембране иммунокомпетентных клеток, включая макрофаги, моноциты, Т- и В- лимфоциты.
Антитела – вещества, синтезируемые плазматическими клетками, способные специфически связываться с введенными в организм антигенами или гаптенами.
Иммунологическая память – способность организма быстро и с большей силой реагировать на повторное введение антигена.
Иммунная система в организме взрослого человека представлена красным костным мозгом – источником стволовых клеток для иммуноцитов, тимусом (центральный орган лимфопоэза), периферическими органами лимфопоэза (селезенка, лимфатические узлы, солитарные и групповые фолликулы в органах) и иммунокомпетентными клетками.
Клетки иммунной системы в функциональном отношении можно разделить на четыре группы (по Климову В.В., 2009):
1. Антигенпрезентирующие клетки (Antigen-presenting cells):
а. Макрофаги, дендритные клетки типов 1 и 2, В-лимфоциты. 2. Регуляторные клетки (Regulatory cells):
а. Естественные регуляторные Т-клетки, Т-индукторы, Т-хелперы типов 1, 2 и 3, Т-регуляторы типа 1.
3. Эффекторные клетки (Effector cells):
72
а. Плазматические клетки (дифференцирующиеся из В- лимфоцитов);
b. Цитотоксические Т-клетки с фенотипом CD+8 (или Т- киллеры);
с. Эффекторные Т-клетки воспаления с фенотипом CD+4 (или Т- лимфоциты, ответственные за гиперчувствительность замедлен-
ного типа);
d. Нейтрофилы, эозинофилы, базофилы, тучные клетки, натуральные киллеры (NK-клетки), макрофаги.
4. Клетки памяти (Memory cells):
а. Т-клетки памяти с фенотипом CD+8, Т-клетки памяти с фенотипом CD+4;
b. Долгоживущие плазматические клетки, В-клетки памяти.
Специфические медиаторы Антитела (АТ)
Это вещества белковой природы, относящиеся к семейству иммуноглобулинов (Ig). АТ синтезируются плазматическими клетками, после чего специфически взаимодействуют с комплементарным антигеном в ходе гуморального иммунного ответа.
Молекула Ig состоит из двух легких цепей (L-цепи) и двух тяжелых (Н-цепи). В цепях различают вариабельную область (V-область) и константную область (С-область). V-область у разных антител варьирует. V-области L и H-цепей образуют антигенсвязывающий центр или Fab-фрагмент – место распознавания или связывания антигена. Константная область молекулы Ig имеет Fc-фрагмент, образованный лишь Н-цепями. Эти области обеспечивают связывание молекулы Ig с клетками-эффекторами (макрофаги, NK-клетки, нейтрофилы, тучные клетки и др.). В результате этого взаимодействия активируются фагоцитоз и цитолиз, развивается дегрануляция тучных клеток.
Различают 5 классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM.
IgG – преобладающий класс АТ, производится в больших количествах при вторичном иммунном ответе и защищает ткани от бактерий, вирусов и токсинов. Циркулирует в крови и лимфе. Fc-фрагмент распознается рецепторами клеток (макрофаги и др.). Соединяется с антигенами и активизирует систему комплемента (опсонизация) и лизис клеток. Он способен проходить через плаценту и из плазмы крови
73
в тканевую жидкость. Содержание повышается при инфекциях, снижается при недостатке гуморального иммунитета.
IgА преобладает в секретах желез (пот, слезы, слизь, молоко, вагинальный, простатический секреты), в слизистых оболочках, в лимфатических узлах и селезенке. Уровень IgА повышается при респираторных и кишечных инфекциях. Это первая линия противомикробной защиты.
IgМ обнаруживается преимущественно внутри сосудов и синтезируется плазмоцитами при первичном попадании антигена в организм. IgМ эффективно агглютинирует бактерии и, кроме того, запускает комплементзависимый цитолиз, осуществляя главную защитную функцию при бактериемии. Не проходит через плаценту (его появление свидетельствует о внутриматочной инфекции).
IgD является маркером В-лимфоцитов, опосредует дифференцировку В-клеток. Не проходит через плаценту, не связывает комплемент.
IgE специфически взаимодействует с тучными клетками и базофилами. При действии аллергенов (пыльца растений, пчелиный яд и др.) IgE связываются с рецепторами Fc-фрагментов IgE, встроенных в клеточную мембрану базофила или тучной клетки, формируя комплекс. При повторном попадании антиген связывается с IgE комплекса, что служит сигналом для дегрануляции. Клинически это проявляется в виде отека, сыпи, приступов астмы, крайняя форма – анафилактический шок. Плазмоциты с IgE находятся в бронхиальных и перитонеальных лимфатических узлах, слизистой оболочке желудочнокишечного тракта, немного в селезенке. Не проходит через плаценту, не связывает комплемент. В крови обнаруживаются лишь следы IgЕ.
Неспецифические медиаторы
К ним относят цитокины, медиаторы гиперчувствительности немедленного типа (гистамин, серотонин, гепарин), интерфероны, лизоцим, а также систему комплемента.
Цитокины
Этим термином принято называть обширное семейство биологически активных пептидов, оказывающих гормоноподобное действие. В зависимости от типа клеток, синтезирующего данный цитокин, различают интерлейкины, монокины и лимфокины. Данные медиаторы обеспечивают взаимодействие клеток иммунной, эндокринной и нервной систем.
74
Интерфероны
Группа белков, продуцируемых вирусинфицированными или активированными клетками. При выделении интерферона в межклеточное пространство он связывается с рецепторами интактных клеток, образуя своеобразный барьер, отграничивающий очаг вирусной инфекции.
Лизоцим
Фермент, расщепляющий мурамиловую кислоту в составе оболочки грамположительных микроорганизмов вплоть до лизиса микробной клетки. Лизоцим синтезируется гранулоцитами, моноцитами и макрофагами. В норме лизоцим присутствует в достаточно высоких количествах в различных физиологических секретах и жидкостях организма.
Комплемент
Представляет группу белков, содержащихся в свежей сыворотке крови человека и животных, активизирующихся при связывании антитела с антигеном и обеспечивающих процесс опсонизации (связывание с антигенами для облегчения их фагоцитоза нейтрофилами).
Виды иммунитета:
неспецифический (врожденный) иммунитет
специфический (приобретенный, адаптационный) иммунитет
Неспецифический (врожденный) иммунитет
Это генетически обусловленное свойство организма обеспечивать свою защиту от любых чужеродных агентов с помощью комплекса факторов. Существуют четыре типа факторов неспецифической защиты: анатомические, физиологические, клеточные, факторы воспаления.
Физические барьеры
К физическим барьерам относятся кожные покровы и слизистые оболочки воздухоносных путей. Кожа играет роль механического барьера, препятствующего проникновению микроорганизмов. А слизистые оболочки дыхательных путей, помимо барьерной функции, обеспечивают удаление и инактивацию патогенов благодаря мерцанию ресничек эпителиоцитов и секрету бокаловидных клеток и желез подслизитой основы.
Физиологические барьеры
Этот тип защиты включает температуру тела, напряженность кислорода, рН, а также растворимые факторы, такие как лизоцим, ин-
75
терферон, комплемент, принимающие участие в естественной невосприимчивости организма к патогенам.
Фагоцитоз
Захват и переваривание микроорганизмов осуществляется «профессиональными» фагоцитами. К ним относятся два типа клеток: макрофаг и нейтрофил. Чтобы осуществился фагоцитоз, должна произойти адгезия микроба на поверхности нейтрофила или макрофага. Это достигается за счет довольно примитивного механизма узнавания, вероятно основанного на взаимодействии углеводных остатков. В зависимости от своей природы микроб, прикрепившийся к поверхностной мембране фагоцита, может инициировать фазу поглощения путем активации актин-миозиновой сократительной системы, что приводит к образованию вокруг микроорганизма псевдоподий. По мере того как близлежащие рецепторы последовательно присоединяются к поверхности микроба, плазматическая мембрана надвигается на микробную клетку подобно застежке-молнии до тех пор, пока она не будет полностью заключена в вакуоль. Далее цитоплазматические гранулы сливаются с фагосомой и изливают в нее свое содержимое, в результате образуется фаголизосома. В ней за счет активности гидролитических ферментов происходит полное или частичное разрушение патогена. Часть разрушенных компонентов микробной клетки удаляется в экстрацеллюлярную среду, другая остается на поверхности фагоцитирующей клетки.
Воспаление
Поврежденные кожные покровы являются входными воротами для проникновения патогенов. Проникшие инфекционные агенты запускают воспалительную реакцию, направленную на удаление микроорганизмов. Наиболее характерные признаки реакции – это увеличение кровоснабжения, которое приводит к избыточному наполнению кровью капиллярной сети и в результате – к локальному покраснению. Повышенная проницаемость капилляров обеспечивает выход в поврежденную ткань плазмы и клеток крови, среди которых наибольшее количество приходится на фагоциты. В формировании воспалительного ответа принимают участие также различные медиаторы. К ним относят медиаторы, выделяемые микроорганизмами, поврежденной тканью, фагоцитирующими клетками, и группу медиаторов плазмы крови, получивших название белков острой фазы. Основным белком этой группы является С-реактивный протеин. Данный белок взаимодействует с полисахаридным компонентом клеточной стенки бактерий и активирует систему комплемента, которая лизиру-
76
ет микроорганизмы или опсонизирует их, способствуя поглощению микроорганизмов фагоцитирующими клетками.
Специфический (приобретенный) иммунитет
Сущность специфического иммунитета отражена уже в самом названии. Ф. Бернет определил иммунитет как реакцию организма, направленную на дифференциацию всего "своего" от всего "чужого". Выделяют два вида приобретенного иммунитета: клеточный и гуморальный.
Клеточный иммунитет
В отличие от гуморального иммунного ответа, где антиген нейтрализуют эффекторные молекулы – антитела, клеточный иммунный ответ опосредован Т-лимфоцитами. Выделяют два основных варианта клеточного иммунного ответа – цитотоксический и воспали-
тельный.
Основными эффекторными клетками цитотоксического клеточного иммунного ответа являются Т-киллеры, которые запрограммированы на синтез рецепторов определенной специфичности и распознают антиген только в комплексе с антигеном гистосовместимости МНС класса I. В процессе распознавания поверхностного антигена цитотоксический Т-лимфоцит вступает в тесный контакт со своей мишенью и уничтожает ее. Процесс лизиса чужеродных клеток состоит из нескольких этапов.
Первый этап – специфическое связывание активированных Т-киллеров с поверхностным чужеродным антигеном (пептидами вирусных трансплантационных, раковых антигенов) в комплексе с МНС класса I. Далее происходит перестройка цитоплазмы Т-киллера, называемая программированием лизиса. В цитоплазме цитотоксического лимфоцита происходит накопление гранул, содержащих растворимые медиаторы, такие как перфорин, цитолизин, гранзимы и др.
Второй этап литического цикла – летальный удар, в ходе которого происходит экзоцитоз медиаторов с образованием трансмембранных пор.
Третий этап, приводящий к лизису клетки-мишени, характеризуется увеличением ее объема за счет все большего проникновения воды через поврежденную мембрану. В результате этих процессов происходят разрыв мембраны клетки-мишени и ее гибель. Эффекторная клетка сохраняется и способна к дальнейшему цитолитическому действию.
Воспалительный тип клеточного иммунного ответа обуслов-
лен функционированием Т-хелперов 1 типа, которые активируют
77
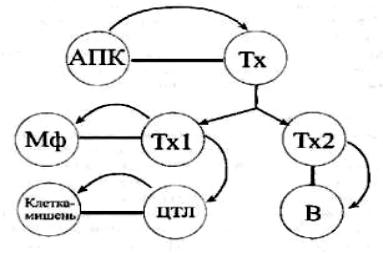
макрофаги, выступающие в роли эффекторных клеток, разрушающих патоген.
Гуморальный иммунитет
Возбудители бактериальных или вирусных заболеваний в процессе своей жизнедеятельности в организме хозяина оказываются в той или иной ситуации во внеклеточной среде, где подвергаются действию антител – эффекторных молекул, продуцируемых В- лимфоцитами.
Гуморальный иммунный ответ осуществляется по принципу
трехклеточной системы кооперации (рис. 29).
На начальных этапах каскада событий проис-
ходит антигенное распо-
знавание, которое в самом начале носит неспецифический характер. Первой клеткой, вступающей во взаимодействие с антигеном, является макрофаг. Антиген может поступать в организм в составе бактериальной клетки или вирусной частицы, а также в виде высокомолекулярного белка или полисахарида, например при вакцинации. И в том и в другом случае он подвергается
фагоцитозу. В результате происходит "разборка" крупных молекул на отдельные "блоки" – процессинг антигена. Такой процессированный антиген экспрессируется на поверхности макрофага в комплексе с собственными антигенами гистосовместимости МНС класса II. Этот комплекс в 100–1000 раз более иммуногенен, чем нативный антиген, и является I специфическим сигналом для распознавания Т- хелперами 2 типа. Одновременно в макрофагах синтезируется интерлейкин I, который секретируется наружу и является II неспецифическим сигналом для распознавания Т-хелперами. В результате происходит двойное распознавание Т-хелпером антигена, т. е. он узнает и чужеродный антиген, и собственный антиген МНС класса II. Хел-
78
перы синтезируют интерлейкины 4, 10, что вызывает пролиферацию и дифференцировку B-клеток. Стимулированные В-лимфоциты трансформируются в плазматические клетки и секретируют Ig G, M. Циркулирующие в крови антитела связываются с антигенными детерминантами поверхностных антигенов бактерий. Комплекс микроба с антителом запускает каскад химических реакций, получивших название классических путей активации комплемента. Конечным продуктом такой активации является образование мембран атакующего комплекса, который наносит клеточной стенке бактерии множественные повреждения. Помимо непосредственного воздействия на бактериальную клетку защитные свойства антител реализуются путем нейтрализации бактериальных токсинов. Молекула антитела присоединяется к токсину вблизи активного центра и стереохимически блокирует его взаимодействие с макромолекулярным субстратом.
Связывание антител с инкапсулированными бактериями облегчает их фагоцитоз лейкоцитами и макрофагами. Этот феномен носит название опсонизации. Как известно, неопсонизированным бактериям, имеющим капсулу, как правило, удается избежать фагоцитоза.
Помимо эффекторных клеток в процессе гуморального иммунного ответа образуются клетки памяти, которые помогают организму противостоять при повторном заражении тем же микроорганизмом. Вторичный иммунный ответ характеризуется более быстрой и эффективной продукцией антител, что является результатом "настройки" антитела образующей системы. Кинетика синтеза иммуноглобулинов разных классов при первичном и вторичном иммунном ответе различна. Так, после первого контакта организма с антигеном в первую очередь удается обнаружить IgM-продуцирующие клетки. Синтез IgG достигает своего максимума лишь на 5–7-е сутки. При вторичном иммунном ответе происходит переключение синтеза иммуноглобулинов с М на G, концентрация которых быстро нарастает, достигая пиковых значений уже на 2-е сутки.
Описанный выше процесс клеточной кооперации подразумевает участие тимусзависимого антигена, для которого взаимодействие хелперных Т-клеток с В-лимфоцитами является обязательным условием развития гуморального иммунного ответа. К тимусзависимым антигенам относятся большинство глобулярных белков, синтетические полипептиды, ксеногенные и аллогенные клетки. Те биополимеры и их синтетические аналоги, которые содержат регулярно повторяющиеся идентичные структуры, способны индуцировать антителообразование без участия Т-хелперов. Такие антигены называют ти-
79
муснезависимыми, в их число входят полисахариды, агрегаты белков регулярного строения и др.
ОПИСАНИЕ МИКРОПРЕПАРАТОВ
1.Мезенхима зародыша цыпленка. Окраска гематоксилином
иэозином (Приложение, рис. 18). На поперечном разрезе зародыша
при малом увеличении микроскопа наблюдается рыхлая сеть связанных друг с другом клеток, заполняющая промежутки между зародышевыми листками. Это малодифференцированная мезенхима. При большом увеличении мезенхимоциты представляются звездчатыми и веретеновидными с округлым или овальным ядром с низкой степенью конденсированности хроматина и слабо базофильной, окрашенной в голубоватый цвет цитоплазмой.
Зарисовать мезенхимные клетки.
2. Кровь лягушки. Мазок. Окраска гематоксилином и эози-
ном (Приложение, рис. 20). При большом увеличении микроскопа эритроциты имеют овальную форму, темное палочковидное ядро и ярко оксифильную цитоплазму. Лейкоциты значительно уступают в численности эритроцитам и подразделяются на гранулоциты и агранулоциты. Тромбоциты представляют собой ядросодержащие клетки веретеновидной формы со слабо окрашенной цитоплазмой и ядром овальной формы.
Зарисовать форменные элементы крови лягушки (эритроциты, лейкоциты, тромбоциты).
3. Кровь человека. Мазок. Окраска азур II – эозином (Прило-
жение, рис. 19). На препарате необходимо найти и зарисовать эритроциты, окрашенные эозином в розовый цвет. Так как эритроциты имеют форму двояковогнутого диска, центральная часть их более тонкая и имеет более светлую окраску. Эритроциты – самые многочисленные клетки крови, и на мазке они составляют большинство. Среди эритроцитов видны лейкоциты (1–5 в поле зрения).
Наиболее часто встречаются сегментоядерные нейтрофилы, имеющие темно-фиолетовое сегментированное ядро (3–5 сегментов) и почти прозрачную (слабо-розовую) цитоплазму с очень мелкой, трудно различимой зернистостью. Эозинофильные гранулоциты имеют слабо базофильную цитоплазму, окрашенную в голубоватый цвет. Цитоплазма заполнена крупными розовыми гранулами одинаковых размеров. Ядро менее плотное, чем у сегментоядерного нейтрофила, обычно имеет два сегмента. Базофильные гранулоциты
80